
Exploring the Bioactive Metabolites with Anti-Malarial Properties
Derived from Endophytic Microbial Resources Indigenous to
Indonesia
Elsera Br Tarigan
1
a
, Partomuan Simanjuntak
2
b
, Dwinna Rahmi
2
c
, Praptiwi
2
d
,
Erwahyuni Endang Prabandari
3
e
, Danang Waluyo
3
f
, Muhammad Ilyas
4
and Andria Agusta
2
g
1
Research Center for Agroindustry, National Research and Innovation Agency Indonesia, Indonesia
2
Research Center for Pharmaceutical Ingredient and Traditional Medicine,
National Research and Innovation Agency Indonesia, Indonesia
3
Research Center for Vaccine and Drug, National Research and Innovation Agency Indonesia, Indonesia
4
Research Center for Biosystematics and Evolution, National Research and Innovation Agency Indonesia, Indonesia
Keywords: Fusarium sp, Hypomycetes, High-Throughput Screening, PfDHODH, Medicinal Plants.
Abstract: Malaria, caused by parasite infection, is still difficult to cure due to partial treatment resistance. This
emphasizes the need for innovative molecules with different modes of action, particularly those derived from
unconventional sources such as endophytic microbes. This study investigates endophytes from medicinal
plants in order to find possible antimalarial drugs that target Plasmodium falciparum dihydroorotate
dehydrogenase (PfDHODH). Endophytes were associated to Mitragyna speciosa, Zingiberaceae, Uncaria
gambir, and others, yielding 586 species. Screening using a 96-well plate produced a z-factor of 0.92,
indicating trustworthy findings. Hypomycetes (from Physalis angulata L.) and Fusarium sp. (from Hornstedtia
scyphifera) showed considerable anti-malarial activity, suppressing PfDHODH by 51.53% and 66.37%,
respectively. Metabolomic profiling using LC-HRMS demonstrated that Hypomycetes included bioactive
substances such as uracil, kojic acid, and phloroglucinol, whereas Fusarium sp.
1 INTRODUCTION
Anopheles mosquitoes carrying the Plasmodium
infection bite humans, transmitting the infectious
disease malaria. The prevalence of malaria in the
world in 2019 was 227 million, and it increased to 241
million people in 2020. Indonesia itself experienced
an increase; in 2013, there were 0.4% of cases, and
this increased in 2018 to 1.4% (Kemenkes 2018).
There are five Plasmodium species that cause malaria
in humans, but Plasmodium falciparum is the most
potent and deadly.
The most effective treatment for malaria infection
is artemisinin-based combination therapy, which has
a
https://orcid.org/0009-0009-8361-1651
b
https://orcid.org/0000-0002-8501-908X
c
https://orcid.org/0000-0002-0987-2345
d
https://orcid.org/0000-0003-1804-4549
e
https://orcid.org/0000-0001-7609-9252
f
https://orcid.org/0000-0003-1280-5984
g
https://orcid.org/0000-0002-9226-6265
no side effects. However, over time, Plasmodium
showed indications of partial resistance to artemisinin
derivatives. Antimalarial drugs can inhibit key
enzymes responsible for cell growth and parasite
development. A group of genes known as P.
falciparum lactate dehydrogenase (PfLDH), P.
falciparum dihydroorotate dehydrogenase
(PfDHODH), and P. falciparum dihydrofolate
reductase (PfDHFR) play a big role in the growth of
plasmodium parasites. PfDHODH is an important
component of parasite metabolism and plays a key
role in the de novo pathway of pyrimidine
biosynthesis. This enzyme facilitates the conversion
of dihydroorotate (DHO) into orotate. This compound
Tarigan, E. B., Simanjuntak, P., Rahmi, D., Agusta, A., Praptiwi, , Prabandari, E. E., Waluyo, D. and Ilyas, M.
Exploring the Bioactive Metabolites with Anti-Malarial Properties Derived from Endophytic Microbial Resources Indigenous to Indonesia.
DOI: 10.5220/0013554600004612
Paper published under CC license (CC BY-NC-ND 4.0)
In Proceedings of BRIN’s 2nd International Conference for Health Research (ICHR 2024), pages 47-53
ISBN: 978-989-758-755-9
Proceedings Copyright © 2025 by SCITEPRESS – Science and Technology Publications, Lda.
47

is essential for generating the pyrimidine bases
required for DNA and RNA synthesis (Akinnusi et al.
2023). However, it is essential to emphasize that
targeted therapy must include safety and specificity,
as these treatments must suppress the parasite while
inflicting minimal harmful effects on the human
body.
Developing antimalarial drugs targeting
PfDHODH is a growing area of research to overcome
malaria drug resistance and improve malaria
treatment. The development of new malaria drugs
begins with the exploration of bioactive plant
metabolites and even plant microorganisms, one of
which is endophyte. Endophytic fungi are
microorganisms that grow in host tissue and produce
bioactive compounds similar to those of their host
without causing disease symptoms. Secondary
metabolites produced by endophytic fungi have many
activities, such as antioxidants and anti-bacterial.
Research on endophytic fungi as a source of anti-
malarial bioactive metabolites is still limited, whereas
Indonesia is rich in germplasm sources.
Research related to the potential of endophyte
bioactive metabolites as antimalarials was carried out
by Ibrahim et al. (2017). The bioactive compound
fusaripeptide found in Fusarium sp., which was
isolated from the roots of the Mentha longifolia L.
plant, potentially kill malaria parasites (Ibrahim et al.,
2017). Fusarium endophytes from Cinchona calisaya
trees could also stop Plasmodium berghei from
growing by 62.18% to 78.41% (Hasbi, 2019).
The objective of this research was to: (1)
Screening of extracts was carried out on 587
endophytic fungi that had PfDHODH inhibitory
activity; (2) up-scaling cultivation and extraction of
Fusarium sp. endophytes. which is associated with
Phyisalus angulata and Hornstedtia scyphifera var.
fusiform, which has a percentage of >50% inhibition
of the PfDHODH enzyme; (3) identified the profile
compounds of extract by using HRMS.
2 METHODS
2.1 Materials
The sample population consisted of 586 endophytes.
Those endophytes was associated with the Mitragyna
speciosa (36); Soultalum album L (15); Phyllanthus
urinaria (4); Myristica fragrans (41); Andrographis
paniculata (18); Piper sarmentosum (36); Physalis
angulata L. (15) ; Chilorantus officinalis (5);
Staurogyne longata (5); Coleus amboinicus (19);
Nigelia sativa (22); Ziziphus mauritiana (50);
Artemisia annua (8); Artemisia vulgaris (13);
Uncaria Gambier (74); Zingiberaceae (95); Piper
nigrum (21); Kaempferia parviflora (26); Murraya
koenigii (4); medicinal plants from Mandalika
Lombok (35); and medicinal plants from Bali (44).
Enzyme inhibition testing was: DMSO, HEPES,
milli-Q water, KOH, NaCl, Triton-X 100, decyl
ubiquinone (d-UQ), absolute ethanol, aluminium foil,
parafilm, L-dihydro orotate (L-DHO),
dichloroindophenol (DCIP), recombinant enzyme
PfDHODH.
2.2 Endophytic Bioproduction and
Extraction
The endophytic fungi were cultivated in potatoes
dextrose broth (PDB) medium. Cultivation was
carried out in large-scale media, 7–10 L, at room
temperature for several weeks until bioactive
metabolites were formed. The process of cultivating
endophytic fungal isolates into PDB media was
carried out sterilely in laminar air flow (LAF). The
extraction was carried out by liquid-liquid extraction
technique using ethyl acetate solvent. The extracted
material was evaporated using a rotary evaporator,
followed by a subsequent drying process with
nitrogen gas. The dried extract was stored at a
freezing temperature for further processing.
2.3 PfDHODH Inhibition Screening
Assay
Measurement refers to the method developed by
Pramisandi et al. (2021), with some modifications.
Screening of extracts/isolates was carried out on 586
endophytic fungi that had PfDHODH inhibitory
activity. The inhibitory activity of key target cells was
assessed using the recombinant enzyme
dihydroorotate dehydrogenase (PfDHODH) derived
from P. falciparum. The inhibition experiments of
PfDHODH were conducted by quantifying the
decrease in the reduction of the electron acceptor 2,6-
dichloroindophenol (DCIP). The 96-well plate was
filled with 2 µL of extract and 190 µL of test solution.
Centrifugation was used to mix the the solutions for
30 seconds at 500 rpm. L-dihydroorotate (L-DHO) 5
mM were added to each well to start the enzymatic
reaction. Inhibitor activity was determined by
spectrophotometer at 600 nm. Inhibitory activity was
calculated based on equation (1).
% Inhibition = 100 −
[
..
.
x 100%]
(1)
ICHR 2024 - BRIN’s International Conference for Health Research (ICHR)
48

The endophyte fungi that had an inhibition percentage
above 50% were selected for bioproduction.
2.4 Compounds Characteristic Using
LC-HRMS
Dried extract (1 mg) was soluble using 1 mL of MS
grade MeOH. Sample analysis was performed
according to Windarsih et al. (2022) with some
modifications. The analysis was conducted using
liquid chromatography with the Thermo Scientific™
Vanquish™ UHPLC Binary Pump, along with
Orbitrap high-resolution mass spectrometry using the
ThermoScientific™ Q Exactive™ Hybrid
Quadrupole-Orbitrap™ High Resolution Mass
Spectrometer. Analytical column of Thermo
Scientific™ Accucore™ Phenyl-Hexyl 100 mm × 2.1
mm ID × 2.6 µm was used for liquid chromatography.
Using a gradient method and a flow rate of 0.3
mL/min, the mobile phases were MS-grade water
containing 0.1% formic acid (A) and MS-grade
methanol containing 0.1% formic acid (B). Initially,
the mobile phase B was set at 5% and then gradually
increased to 90% over a period of 16 minutes. After
that, it remained at 90% for 4 minutes and then
returned to the initial condition (5% B) until 25
minutes. The temperature of the column was adjusted
to 40 ◦C, while the injection volume was set at 3 µL.
An untargeted screening was conducted using full
MS/dd-MS2 acquisition mode in either positive or
negative ionization polarities/states. The spray
voltage was set to 3.30 kV, while the capillary
temperature and the auxiliary gas heater temperature
were set at 320 ◦C and 30 ◦C, respectively. The scan
range was conducted from 66.7 to 1000 m/z, with a
resolution of 70,000 for full MS and 17,500 for dd-
MS2, in both positive and negative ionisation modes.
3 RESULT AND DISCUSSION
3.1 Enzymatic Screening of Endophyte
Extract as an Anti-Malarial
Endophytic fungi are a type of fungus that live in
plant tissue without causing visible damage or disease
in their hosts (Ababutain et al. 2021). The metabolites
produced are generally similar as those of the host
plant. Metabolites generated by endophytes exhibit
numerous functions, including antimalarial activity.
Malarial assay can be carried out enzymatically,
parasitically and computationally. PfDHODH, P.
falciparum Dihydroorotate Dehydrogenase, plays a
vital role as an enzyme in the pyrimidine biosynthesis
pathway of the malaria parasite P. falciparum.
Primary screening was conducted on 586
endophyte extracts derived from a variety of plant
categories and parts, such as leaves, stems, roots, and
seeds. Those extracts were plated on a 96-well plate
for the PfDHODH assay, Fig.1. There are numerous
critical stages that comprise its mechanism. The
enzyme attaches to its substrate, dihydroorotate, at its
active site and facilitates the chemical reaction that
converts dihydroorotate into orotate. This procedure
entails the extraction of two electrons and two protons
from dihydroorotate. PfDHODH utilizes the
coenzyme FMN (Flavin Mononucleotide) as an
electron acceptor, facilitating the conversion of FMN
from its oxidized state (blue form) to its reduced state
(red color), Fig.2. FMN transports electrons to the
electron transport chain located in the mitochondrial
membrane of P. falciparum. This process produces
vital energy for the parasite's survival and growth.
The end product of the reaction, orotate, is liberated
from the active site of the enzyme, so concluding the
conversion process. PfDHODH plays a crucial role in
the formation of pyrimidine, making it an important
target for the development of antimalarial drugs.
Inhibiting this enzyme can effectively hinder the
growth and reproduction of P. falciparum (A. Phillips
dan K. Rathod 2012). PfDHODH facilitates an
enzymatic process that is linked to the reduction of
DCIP. PfDHODH transfers electrons to DCIP,
converting it from its oxidized blue form (DCIPox) to
its reduced colorless form (DCIPred), as it converts
dihydroorotate to orotate and reduces
decylubiquinone to decylubiquinol, Fig. 2.
During this study, extracts that demonstrated
inhibitory efficacy exceeding 50% were designated as
primary findings. There were two extracts identified
as the main hits (blue color), particulary endophytes
associated with Physalis angulata L (51,53%),
Hornstedtia scyphifera (66,37%). The two
endophytes were fungi belonging to the Hypomycetes
type, which was associated with Physalis angulata L,
and the Fusarium sp., which was associated with
Hornstedtia scyphifera. The batch codes for these
fungi were BgPa1 and HSFP3, respectively.
Exploring the Bioactive Metabolites with Anti-Malarial Properties Derived from Endophytic Microbial Resources Indigenous to Indonesia
49
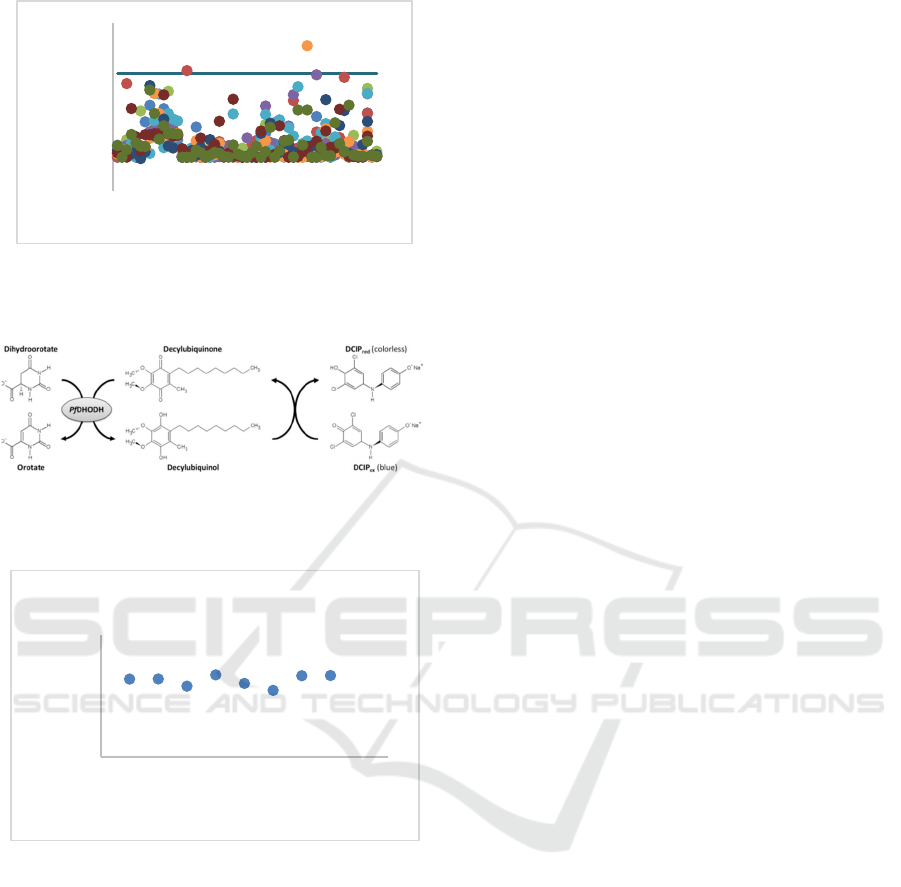
Figure 1: Inhibitory activity of 586 microbial extracts
against PfDHODH; purple-dashed line represents threshold
line.
Figure 2: Endophytes extracts are subjected to enzyme-
based screening assays to determine their inhibitory
activities against PfDHODH (Waluyo et al. 2021)
Figure 3: The z-factor value of each assay batch.
In this HTS study the z score value was also
measured. The Z'-factor is a statistical metric that
quantifies the reliability of data acquired from high-
throughput screening (HTS) experiments. A high Z'-
factor indicates that the assay is capable of accurately
differentiating between active and inactive
substances. The mean value of the z'factor in this
research was 0.92 (Figure 3), indicating excellent
results.
3.2 Untargeted Compounds Assay
Liquid chromatography-high resolution mass
spectrometry (LC-HRMS) was employed for
untargeted compounds, allowing for a comprehensive
screening of small compounds in the extract. The
most often used approach for analysis is reversed-
phase liquid chromatography (RPLC), specifically
the C18-based method. LCHRMS enables the precise
identification of a comprehensive metabolomic
profile, encompassing both major and minor
components in natural extracts. The novel
information obtained from LC/HRMS analysis of the
metabolomic profile of endophytes Hypomycetes
(associated with Physalis angulata) and Fusarium sp
(associated with Hornstedtia scyphifera) can be
useful for advancing study. Results of the
chromatogram analysis for each extract using
LC/HRMS are displayed in Figure 1.
The LCHRMS results for the ethyl acetate extracts of
endophytes associated with Physalis angulate and
Hornstedtia scyphifera are displayed in Table 1. The
three compounds with the largest areas for
Hyphomycetes endophyte extracts were uracil,
hexadecane, and 3-[(4-hydroxyphenyl)methyl]-
octahydropyrrole[1,2-a]pyrazine-1,4-dione. The
molecular structure of this chemical is complex, and
it exhibits antifungal, anticancer, and anti-
inflammatory properties, respectively (Zhang et al.
2013; Bao et al. 2023; Husnawati et al. 2023). A
fungus belonging to the Hyphomycetes class was
found in the flower of PA. This fungus showed both
antioxidant and antibacterial properties (Palupi et al.
2021). Regarding Fusarium sp. fungus isolates
related with the Hornstedtia scyphifera plant, the
three predominant chemicals were amide compounds,
specifically docosanamide, stearamide, and
hexadecanamide. According to research conducted
by (Gurning 2024) amide compounds exhibit
potential as antidiabetic. Research related to the
potential of endophyte bioactive metabolites as
antimalarials was carried out by Ibrahim et al. (2018).
The bioactive compound fusaripeptide found in
Fusarium sp., which was isolated from the roots of
the Mentha longifolia L, can kill malaria parasites
(Ibrahim et al., 2017). Fusarium endophytes from
Cinchona calisaya trees could also stop Plasmodium
berghei from growing by 62.18% to 78.41% (Hasbi
2019) Considering that Indonesia is known as a
source of germplasm with high biodiversity, it is
certainly a great opportunity to explore endophytic
bioactive metabolites as anti-malarials.
-20
0
20
40
60
80
Inhibition (%)
Number of Extract
0,3
0,5
0,7
0,9
1,1
1,3
0510
Z' Factor Value
Number of screening
Average Z' Factor: 0,92
ICHR 2024 - BRIN’s International Conference for Health Research (ICHR)
50
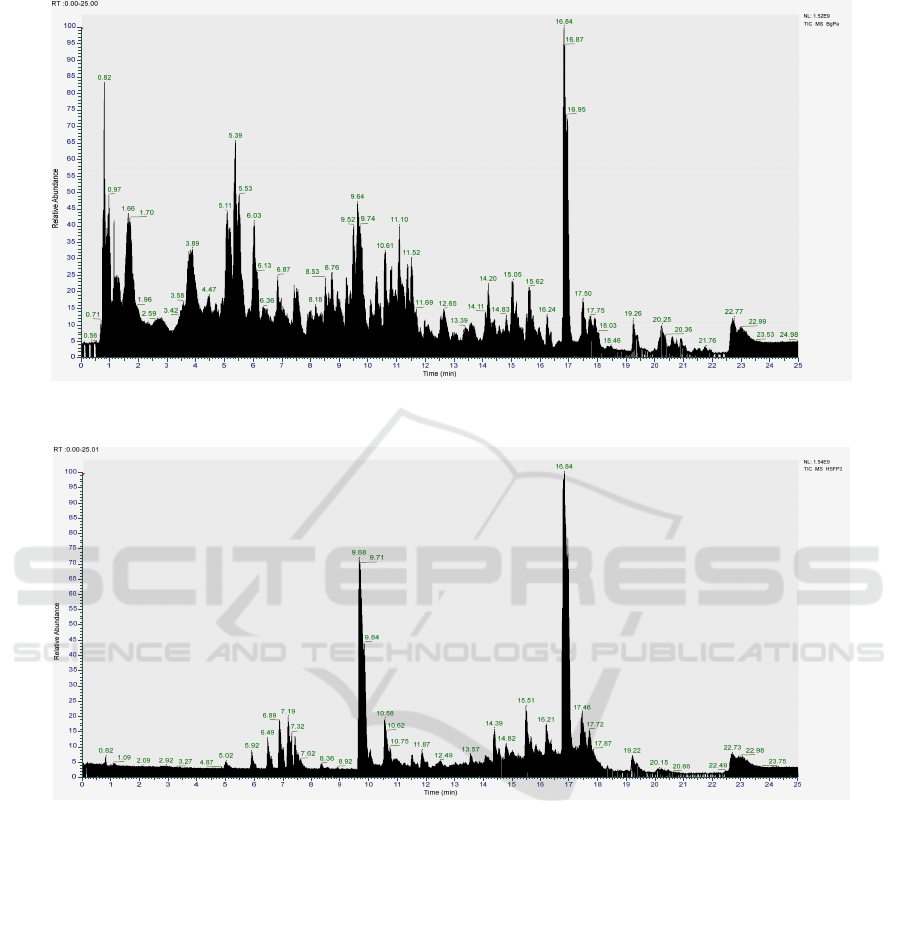
Figure 4: Profile LC/HRMS of endophyte extract: (a) Hypomycetes of endophytes associated with Physalis angulate (b)
Fusarium sp. endophytes associated with Hornstedtia scyphifera.
4 CONCLUSIONS
There were two extracts had an inhibitory PfDHODH
of more than 50%, from endophytes associated to
Physalis angulata L (51.53%) and Hornstedtia
scyphifera (66.37%). These endophytes were
identified as Hypomycetes and Fusarium sp.
Analysis of untargeted compounds using LC-
HRMS on ethyl acetate extracts from endophytes
related to Physalis angulata and Hornstedtia
scyphifera revealed various metabolites with
potential biological activity. This research shows that
endophytes have significant potential as a source of
bioactive metabolites for the development of
antimalarial drugs. Especially considering
Indonesia's high biodiversity, further exploration of
endophyte bioactive metabolites as antimalarials is
very promising.
a
b
Exploring the Bioactive Metabolites with Anti-Malarial Properties Derived from Endophytic Microbial Resources Indigenous to Indonesia
51
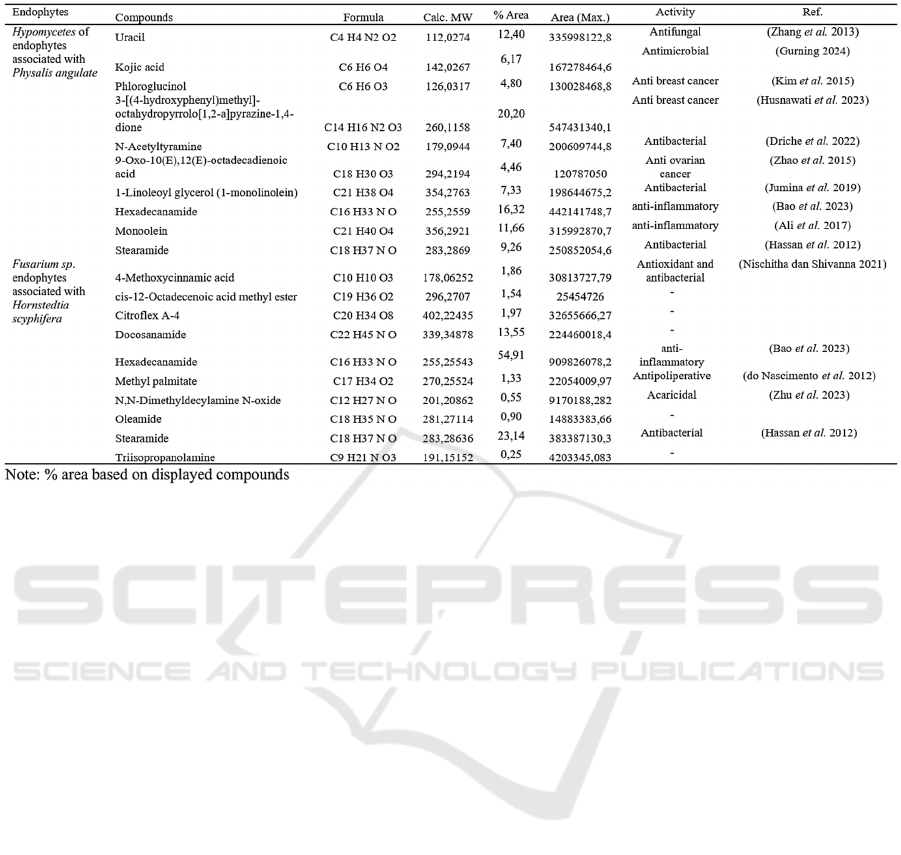
Table 1: Metabolomics components with the highest peak area in the etyl acetate extract.
REFERENCES
A. Phillips M, K. Rathod P. 2012. Plasmodium
Dihydroorotate Dehydrogenase: A Promising Target
for Novel Anti-Malarial Chemotherapy. Infect Disord -
Drug Targets. 10(3):226–239. doi:10.2174/187
152610791163336.
Ababutain IM, Aldosary SK, Aljuraifani AA, Alghamdi AI,
Alabdalall AH, Al-Khaldi EM, Aldakeel SA, Almandil
NB, Abdulazeez S, Borgio JF. 2021. Identification and
Antibacterial Characterization of Endophytic Fungi
from Artemisia sieberi. Int J Microbiol. 2021.
doi:10.1155/2021/6651020.
Akinnusi PA, Olubode SO, Adebesin AO, Osadipe TJ,
Nwankwo DO, Adebisi AD, Titilayo I BA, Alo YM,
Owoloye A, Oyebola KM. 2023. Structure-based
scoring of anthocyanins and molecular modeling of
PfLDH, PfDHODH, and PfDHFR reveal novel
potential P. falciparum inhibitors. Informatics Med
Unlocked. 38 March:101206. doi:10.1016/j.
imu.2023.101206.
Ali I, Manzoor Z, Koo JE, Moon SR, Byeon SH, Yoo ES,
Kang HK, Hyun JW, Lee NH, Koh YS. 2017.
Monoolein, isolated from Ishige sinicola, inhibits
lipopolysaccharide-induced inflammatory response by
attenuating mitogen-activated protein kinase and NF-
κB pathways. Food Sci Biotechnol. 26(2):507–511.
doi:10.1007/s10068-017-0070-x.
Bao L, Sun H, Zhao Y, Feng L, Wu K, Shang S, Xu J, Shan
R, Duan S, Qiu M, et al. 2023. Hexadecanamide
alleviates Staphylococcus aureus-induced mastitis in
mice by inhibiting inflammatory responses and
restoring blood-milk barrier integrity. PLoS Pathog. 19
11 November:1–22. doi:10.1371/journal.ppat.1011764.
Driche EH, Badji B, Bijani C, Belghit S, Pont F, Mathieu
F, Zitouni A. 2022. A New Saharan Strain of
Streptomyces sp. GSB-11 Produces Maculosin and N-
acetyltyramine Active Against Multidrug-Resistant
Pathogenic Bacteria. Curr Microbiol. 79(10):1–10.
doi:10.1007/s00284-022-02994-3.
Gurning K. 2024. Untargeted LC / HRMS Metabolomics
Anal y sis and Anticancer Activit y Assa y on MCF-7
and A549 Cells from Coleus amboinicus Lour Leaf E x
tract. 23 December:1–8.
Hasbi N. 2019. Aktivitas filtrat fusarium endofit asal pohon
kina (Cinchona calisaya ) sebagai anti-plasmodium
nurmi hasbi.
Hassan WHB, Al-Oqail M, Ahmad MS, Al-Rehaily AJ.
2012. Bioactive long chain amides from Solanum
schimperianum hochst growing in Saudi Arabia. Biosci
Biotechnol Res Asia. 9(2):593–599. doi:10.13005/bbr
a/1037.
Husnawati, Kusmardi K, Kurniasih R, Hasan AZ,
Andrianto D, Julistiono H, Priosoeryanto BP, Artika
IM, Salleh MN. 2023. Investigation of Chemical
Compounds from Phomopsis Extract as Anti-Breast
Cancer Using LC-MS/MS Analysis, Molecular
Docking, and Molecular Dynamic Simulations. Int J
Technol. 14(7):1476–1486. doi:10.14716/ijtech.v
14i7.6696.
Ibrahim SRM, Abdallah HM, Elkhayat ES, Al Musayeib
NM, Asfour HZ, Zayed MF, Mohamed GA. 2018.
Fusaripeptide A: new antifungal and anti-malarial
cyclodepsipeptide from the endophytic fungus
ICHR 2024 - BRIN’s International Conference for Health Research (ICHR)
52

Fusarium sp. J Asian Nat Prod Res. 20(1):75–85.
doi:10.1080/10286020.2017.1320989.
Jumina J, Lavendi W, Singgih T, Triono S, Steven
Kurniawan Y, Koketsu M. 2019. Preparation of
Monoacylglycerol Derivatives from Indonesian Edible
Oil and Their Antimicrobial Assay against
Staphylococcus aureus and Escherichia coli. Sci Rep.
9(1):1–8. doi:10.1038/s41598-019-47373-4.
Kemenkes. 2018. Hasil utama Riskesdas 2018. Jakarta.
Kim RK, Uddin N, Hyun JW, Kim C, Suh Y, Lee SJ. 2015.
Novel anticancer activity of phloroglucinol against
breast cancer stem-like cells. Toxicol Appl Pharmacol.
286(3):143–150. doi:10.1016/J.TAAP.2015.03.026.
do Nascimento AM, Conti R, Turatti ICC, Cavalcanti BC,
Costa-Lotufo L V., Pessoa C, de Moraes MO, Manfrim
V, Toledo JS, Cruz AK, et al. 2012. Bioactive extracts
and chemical constituents of two endophytic strains of
Fusarium oxysporum. Rev Bras Farmacogn.
22(6):1276–1281. doi:10.1590/S0102-695X201200
5000106.
Nischitha R, Shivanna MB. 2021. Metabolite
fingerprinting, in vitro antimicrobial and antioxidant
activities and in-silico docking in Alloteropsis cimicina
and its endophytic fungus Penicillium pinophilum. Mol
Biol Rep. 48(5):4021–4037. doi:10.1007/s11033-021-
06410-0.
Palupi KD, Ilyas M, Agusta A. 2021. Endophytic fungi
inhabiting Physalis angulata L. plant: Diversity,
antioxidant, and antibacterial activities of their ethyl
acetate extracts. J Basic Clin Physiol Pharmacol.
32(4):823–829. doi:10.1515/jbcpp-2020-0479.
Pramisandi A, Kurnia K, Chrisnayanti E, Bernawati P,
Dobashi K, Mori M, Mahsunah AH, Nonaka K,
Matsumoto A, Kristiningrum, et al. 2021. Gentisyl
alcohol and homogentisic acid: Plasmodium falciparum
dihydroorotate dehydrogenase inhibitors isolated from
fungi. J Gen Appl Microbiol. 67(3):114–117.
doi:10.2323/jgam.2020.08.004.
Waluyo D, Prabandari EE, Pramisandi A, Hidayati DN,
Chrisnayanti E, Puspitasari DJ, Dewi D, Suryani,
Kristiningrum, Oktaviani AN, et al. 2021. Exploring
natural microbial resources for the discovery of anti-
malarial compounds. Parasitol Int. 85:102432.
doi:10.1016/j.parint.2021.102432.
Windarsih A, Suratno, Warmiko HD, Indrianingsih AW,
Rohman A, Ulumuddin YI. 2022. Untargeted
metabolomics and proteomics approach using liquid
chromatography-Orbitrap high resolution mass
spectrometry to detect pork adulteration in Pangasius
hypopthalmus meat. Food Chem. 386 July
2021:132856. doi:10.1016/j.foodchem.2022.132856.
Zhang G, Zhang Y, Qin J, Qu X, Liu J, Li X, Pan H. 2013.
Antifungal Metabolites Produced by Chaetomium
globosum No.04, an Endophytic Fungus Isolated from
Ginkgo biloba. Indian J Microbiol. 53(2):175–180.
doi:10.1007/s12088-013-0362-7.
Zhao B, Tomoda Y, Mizukami H, Makino T. 2015. 9-Oxo-
(10E,12E)-octadecadienoic acid, a cytotoxic fatty acid
ketodiene isolated from eggplant calyx, induces
apoptosis in human ovarian cancer (HRA) cells. J Nat
Med
. 69(3):296–302. doi:10.1007/s11418-015-0892-x.
Zhu K, Liu X, Qi X, Liu Q, Wang B, Sun W, Pan B. 2023.
Acaricidal activity of bioactive compounds isolated
from Aspergillus oryzae against poultry red mites,
Dermanyssus gallinae (Acari: Dermanyssidae). Vet
Parasitol. 320 July:109983. doi:10.1016/j.v
etpar.2023.109983.
Exploring the Bioactive Metabolites with Anti-Malarial Properties Derived from Endophytic Microbial Resources Indigenous to Indonesia
53
