
Stereo-Event-Camera-Technique for Insect Monitoring
Regina Pohle-Fr
¨
ohlich
a
, Colin Gebler and Tobias Bolten
b
Institute for Pattern Recognition, Niederrhein University of Applied Sciences, Krefeld, Germany
Keywords:
Event Camera, Segmentation, Insect Monitoring, Depth Estimation.
Abstract:
To investigate the causes of declining insect populations, a monitoring system is needed that automatically
records insect activity and additional environmental factors over an extended period of time. For this reason,
we use a sensor-based method with two event cameras. In this paper, we describe the system, the view volume
that can be recorded with it, and a database used for insect detection. We also present the individual steps
of our developed processing pipeline for insect monitoring. For the extraction of insect trajectories, a U-Net
based segmentation was tested. For this purpose, the events within a time period of 50 ms were transformed
into a frame representation using four different encoding types. The tested histogram encoding achieved the
best results with an F1 score for insect segmentation of 0.897 and 0.967 for plant movement and noise parts.
The detected trajectories were then transformed into a 4D representation, including depth, and visualized.
1 INTRODUCTION
Climate and human-induced landscape changes have
a major impact on biodiversity. One process that has
been observed and scientifically documented in recent
years is the decline of many insect species (Hallmann
et al., 2017). To better understand the causes, biodi-
versity monitoring at the species level is needed, but is
currently hampered by several barriers (W
¨
agele et al.,
2022). For example, automated species identifica-
tion is difficult because only a limited number of test
datasets for AI-based techniques for the various mon-
itoring methods are currently available (Pellegrino
et al., 2022). On the other hand, manual monitoring of
insects is costly, which means that often only small ar-
eas and limited time periods are surveyed. Due to the
high cost, such monitoring is only done on a random
basis, so that comparisons for the same habitat type
between regions or between different time periods are
not possible. In addition, manual observations may be
subject to unintentional bias (Dennis et al., 2006) and
their quality may depend on the expertise of the ob-
server (Sutherland et al., 2015). Furthermore, because
the observational sources in this case e.g. as images
of the measurements are not preserved, the accuracy
of the data may be questioned later. This is especially
true because human attention is low for non-foveal
a
https://orcid.org/0000-0002-4655-6851
b
https://orcid.org/0000-0001-5504-8472
visual tasks and for moving objects (Ratnayake et al.,
2021).
Another problem is that even many automated
methods (traps, camera-based systems) can only sur-
vey very small areas. The data thus obtained are
not easily extrapolated to larger areas. In addition,
species interactions or changes within populations are
often difficult to detect due to the limited time win-
dow considered or the size of the observation patch.
Suitable technologies for large-scale and long-term
automated biodiversity monitoring are still lacking
(W
¨
agele et al., 2022).
For this reason, we want to develop a new mon-
itoring method using event cameras. The operation
and output paradigm of this sensor is fundamentally
different from conventional cameras. For example,
the event camera does not capture images at a fixed
sampling rate, but generates event streams (Gallego
et al., 2020). For each detected brightness change
at a pixel position above a defined threshold, the x-
and y-coordinate, a very precise time stamp t in mi-
croseconds and an indicator p for the direction of the
brightness change are recorded. Other advantages
compared to conventional CCD/CMOS sensors in-
clude higher dynamic range, lower power consump-
tion, smaller data volume, and much higher temporal
resolution. Since each pixel of an event camera oper-
ate independently and asynchronously based on rela-
tive brightness changes in the scene, they can also be
used under difficult lighting conditions (strong bright-
Pohle-Fröhlich, R., Gebler, C. and Bolten, T.
Stereo-Event-Camera-Technique for Insect Monitoring.
DOI: 10.5220/0012326500003660
Paper published under CC license (CC BY-NC-ND 4.0)
In Proceedings of the 19th International Joint Conference on Computer Vision, Imaging and Computer Graphics Theory and Applications (VISIGRAPP 2024) - Volume 3: VISAPP, pages
375-384
ISBN: 978-989-758-679-8; ISSN: 2184-4321
Proceedings Copyright © 2024 by SCITEPRESS – Science and Technology Publications, Lda.
375

Figure 1: Position of a bee (white dots) during takeoff in
eight frames captured at 30 frames per second.
ness differences in the scene). First tests with this sen-
sor for insect monitoring have already been success-
fully performed (Pohle-Fr
¨
ohlich and Bolten, 2023).
In this paper we want to discuss further develop-
ments of this approach. These major contributions are
• the use of a stereo event camera setup, so that 4D
data points (x, y, z, t) can be recorded
• the investigation of different event encoding types
for trajectory extraction
• the description of the depth calculation pipeline.
The rest of this paper is structured as follows. Sec-
tion 2 gives an overview of related work. Section 3
describes the dataset acquisition setup. In Section 4,
the currently used processing pipeline is explained
and in Section 5, the obtained results are presented
and discussed. Finally, a short summary and an out-
look on future work is given.
2 RELATED WORK
The use of AI methods has significantly improved in-
sect monitoring in recent years. Although the use of
traps is still a common method for determining the
biomass and abundance of individual insect species,
whereas in the past traps were mostly evaluated by
humans, this is now being done in part with the
help of DNA metabarcoding (Pellegrino et al., 2022)
or automated camera systems. The cameras cap-
ture single images of specific locations where insects
are attracted by targeted lighting, colored stickers,
or pheromones in order to identify and count them
((Qing et al., 2020), (Marcasan et al., 2022), (Dong
et al., 2022)). During image acquisition, the insects
hardly move and can be well segmented due to the
uniform background color used at these locations.
To detect interactions between different insect
species, video cameras are usually used to examine
small areas with very few plants. Under these condi-
tions, good detection results can be obtained for some
insects, such as bees ((Bjerge et al., 2022), (Droissart
et al., 2021)). However, problems arise with trajec-
tory detection (Figure 1), since required high tem-
poral resolution cannot be used due to the required
storage space. In addition, image compression (Fig-
ure 2) is required for long-term monitoring for the
same reason, which makes insect detection difficult
and error-prone.
The monitoring method proposed in this paper
uses event cameras. Various approaches for segmen-
tation and classification of event data can be found
in the literature (Gallego et al., 2020). A common
method for data analysis is to convert the event stream
into 2D images. In this process, all events within
a time window of fixed length or a fixed number of
events are projected into an image using different en-
coding methods preserving different amounts of tem-
poral information (e.g., binary encoding, linear time-
surface encoding, polarity encoding). Then, different
neural networks (e.g., U-Net, Mask-RCNN) are used
for segmentation. In addition, neural networks that
take a point cloud as input (PointNet++, LSA-Net, A-
CNN) are used for both segmentation and classifica-
tion ((Bolten et al., 2022a), (Bolten et al., 2023b)).
3 DATASET RECORDING SETUP
3.1 Used Sensor System
Contrary to the work of (Bolten et al., 2023b) on hu-
man activity recognition, the motion of insects does
not take place on planes, but in 3D space. For this
reason, capturing the scene with only one camera to
estimate the z-coordinate as in (Bolten et al., 2022b)
Figure 2: Poor visibility of a honeybee (see arrow) due to
H.264 video compression.
VISAPP 2024 - 19th International Conference on Computer Vision Theory and Applications
376

Figure 3: Stereo system with two event cameras.
is not sufficient. Therefore, a stereo system was cho-
sen for image acquisition. By capturing the depth
information, it is also possible to make assumptions
about the insect’s size and flight speed and use them
to group the insects. Figure 3 shows the setup of our
measurement system.
Two parallel event camera IMX636 HD sensors
distributed by Prophesee (Prophesee, 2023b) with an
image resolution of 1280 x 720 pixels, each equipped
with an 5mm wide-angle lens, are synchronized in
time and connected to a RaspberryPi 4B running the
data acquisition software. The event stream gener-
ated by each camera is written to an external hard
drive. Additionally, a temperature sensor is integrated
into the measurement setup. The camera system is
powered by a portable power station, which can be
recharged by a solar panel if required, to enable mea-
surements in any terrain. Figure 4 shows our mea-
surement system in action. For manual logging and
labeling of insect activity, an additional event camera
was connected to a laptop in order to display the ref-
erence data.
3.2 Covered View Volume
In order to obtain information about the detection
range of our monitoring system, it was first deter-
mined by calculation. Subsequently, the theoretically
Figure 4: System in action.
Figure 5: View volume.
calculated values were evaluated by throwing refer-
ence objects similar in size to certain groups of in-
sects at a defined distance into the field of view of the
camera system. The measured values confirmed our
calculations. Figure 5 shows the obtained view vol-
ume. The black numbers show the calculated values
and the red numbers show the experimentally deter-
mined values.
Insects with a size of 5 x 2 mm², which is approxi-
mately the size of a mosquito, can be reliably detected
up to a distance of 2 m from the camera. The area
over which these insects can be reliably recognized
is 2.9 m² considered in the top view. Insects with a
size of 10 x 5 mm², which corresponds to the size of
a house fly, can be reliably identified up to a distance
of 5 m from the camera. These insects can be located
on an covered ground area of 18 m². Insects with a
size of 15 x 10 mm², which corresponds to the size
of a honey bee, can still be reliably recognized at a
distance of 7 m from the camera. In this case, the
covered ground area is 35 m². This is a significant in-
crease compared to the ground area of 1 m² used in
manual insect counting.
3.3 Used Database
For the detection of insect flight paths, on the one
hand, the labeled dataset from (Pohle-Fr
¨
ohlich and
Bolten, 2023) was used, which was, however, only
recorded with one camera and not as stereo data. On
the other hand, additional monocular event data were
recorded and labeled. These recordings were taken in
the evening in a garden with grass, and in the morning
and in the afternoon on a balcony with flowers in bal-
cony boxes. Because of the different scenarios, plant
movement and other environmental factors varied.
All files were labeled to include two classes: In-
sect trajectory events and events due to noise and
plant movements. Figure 6 shows a section of one
of the labeled datasets, with the insect class events
colored red and the environment class events colored
white. Thus, eight recordings with a total dataset
Stereo-Event-Camera-Technique for Insect Monitoring
377
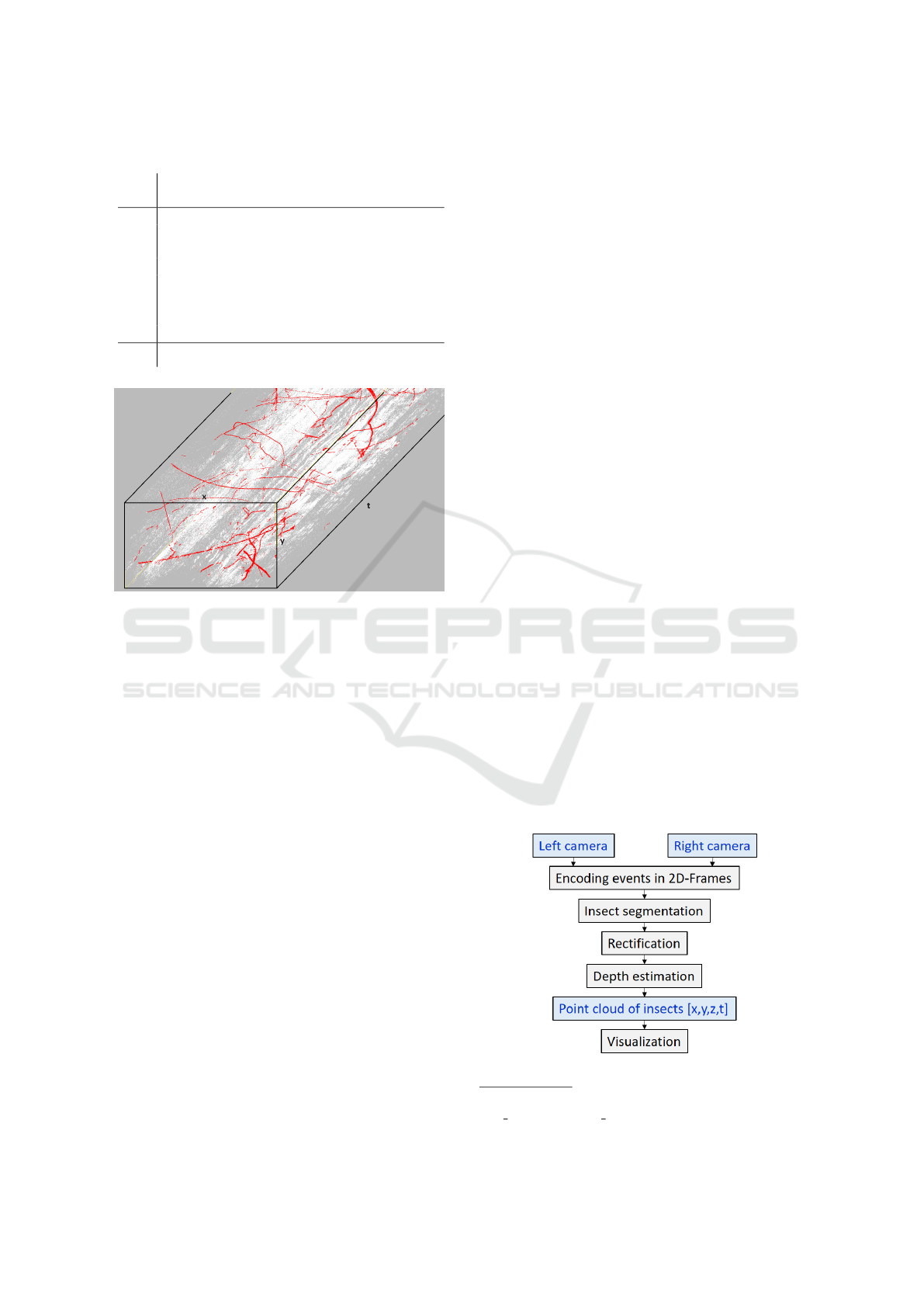
Table 1: Structure of the dataset.
No. Content Size No. of No. of
in sec insect events other events
1 Meadow 30.94 834 373 1 109 131
2 Garden 100.01 1 054 489 200 102
3 Garden 97.40 1 022 401 137 577
4 Garden 156.58 935 939 545 659
5 Balcony 73.24 1 407 740 656 936
6 Balcony 61.12 160 504 756 339
7 Balcony 16.10 78 832 1 037 434
8 Balcony 78.85 1 407 740 656 936
614.24 6 902 018 5 100 114
Figure 6: Example of the labeled insect dataset No. 1
(Events from flight paths are red and events from environ-
mental effects are colored white.).
length of 10 minutes and 14 seconds were available
for neural network training. The basic properties of
the dataset are shown in Table 1. It can be seen that
the plant events are currently still slightly underrepre-
sented.
4 FLIGHT PATH
SEGMENTATION
4.1 Software Pipeline
The 4D trajectory determination for the individual in-
sects is realized in several steps. After recording the
event streams with the two cameras, they are first en-
coded into 2D frames for segmentation with a CNN.
The events classified as insects are then transformed
into rectified coordinates before the depth is calcu-
lated from the disparity of the pixels in each of two
corresponding projected views from the left and right
camera. Finally, the 4D points available after this step
are visualized. The process is illustrated in Figure 7.
The individual steps are described in more detail be-
low.
4.2 Event Encoding
There are a variety of methods for encoding and pro-
cessing the output stream of an event camera (Gal-
lego et al., 2020). Often, the events are converted into
classical 2D images and then processed using estab-
lished image processing methods. This approach was
also chosen for the insect data. Direct processing as
a point cloud would require a reduction in the num-
ber of events by applying downsampling methods.
However, since the insect trajectories represent very
fine structures within the point cloud, these would be
lost. Splitting the output data into different window
regions, as in (Bolten et al., 2023a), would also be
problematic because insects move very fast and thus
fly over several windows in a very short time. The
data would then have to be reassembled for final anal-
ysis. Since time information is lost when event data is
projected into a 2D binary or polarity representation,
various encoding methods have been described in the
literature to preserve this information. Four different
encoding approaches have been investigated for de-
tection of flight trajectories.
• Histogram Encoding
A simple way of encoding is to create a his-
togram. For a given time window, the number of
events per pixel position is determined separately
for each polarity, with the maximum number set
to 5 (Prophesee, 2023a)
1
. This results in a two-
channel encoding. Fast moving objects, such as
insects, have lower frequencies compared to slow
and cyclically moving objects, such as plants. In
the experiments performed, we used a time win-
dow of 50 ms for event accumulation.
• Linear Timesurface Encoding
The linear time surface is another method of en-
coding the time information of events in a 2D rep-
Figure 7: Software pipeline for detection of flight paths.
1
https://docs.prophesee.ai/stable/tutorials/ml/
data processing/event preprocessing.html
VISAPP 2024 - 19th International Conference on Computer Vision Theory and Applications
378

resentation. In this method, the scaled time stamp
of the last event that occurred is stored in each
pixel, again considering the polarities separately.
This yields to a two-channel encoding. The scal-
ing is linear in the range between 0 and 255. In our
experiments we used a time window of 50 ms.
• Event Cube Encoding
Event cubes were developed to combine the infor-
mation from the histogram with the time informa-
tion from the linear timesurface. In event cubes,
each time bin is further divided into three micro
time bins. Instead of counting an event based on
its position and time stamp as in the histogram, the
event is counted linearly weighted according to its
time distance from the center of the neighboring
micro time bins (Prophesee, 2023a)
2
. Since the
summation is done separately for each polarity.
This results in a six-channel encoding and leading
to a higher computational complexity for training
and inference of neural networks compared to the
other encodings. To obtain enough events in a
frame for the calculation, a time window of 50 ms
was used.
• Optical Flow Encoding
Finally, to encode the temporal information con-
tained in the event stream, the optical flow was
also examined, since insects usually move very
fast and in arbitrary directions, while plants move-
ments are slower and occur only in certain direc-
tions.
There are several ways to estimate optical flow.
The algorithm provided in the Metavision SDK
for computing sparse optical flow has the disad-
vantage that clustering is performed beforehand
(Prophesee, 2023a)
3
. Experiments have shown
that, for insect movements in the background with
very few points in the projected image plane, no
optical flow value is calculated.
For this reason, the dense optical flow algorithm
provided by the Metavision SDK was used as
well. Since the data from the insect crossings con-
tains very few objects within a short time window,
training the neural network used for the dense
flow calculation did not produce satisfactory re-
sults, so we used a pre-trained network with the
weights provided in the SDK for our calculations.
As with the other methods, the time window for
optical flow calculation was 50 ms. From the two
predicted components of the flow vector, we have
2
https://docs.prophesee.ai/stable/tutorials/ml/
data processing/event preprocessing.html
3
https://docs.prophesee.ai/stable/samples/modules/cv/
sparse flow cpp.html
Figure 8: Example for optical flow prediction for three dif-
ferent moving objects. The adjacent color wheel was used
for display. The intensity indicates the magnitude of the
flow and the color the direction of the movement.
calculated the magnitude from this estimated ve-
locity vector and direction as the angle to x-axis
between 0 and 360 degrees. Again, resulting in
a two-component encoding. It should be noted,
however, that the computational cost of determin-
ing the optical flow is high. An example of the
predicted optical flow can be found in Figure 8.
In our current investigations, all events in our dataset
were encoded using the four encoding methods, re-
sulting in a total number of 12230 images of each en-
coding.
4.3 Insect Segmentation
After frame encoding, classical 2D networks can be
used for semantic segmentation of trajectories. A
typical network that can perform segmentation with
a relatively small amount of input data is the U-Net
(Ronneberger et al., 2015). It is an encoder-decoder
network with a typical symmetric structure. The en-
coder part is used for feature extraction, where the
convolutional and pooling layers also lead to down-
sampling and information concentration, but also to
spatial mapping reduction. Upsampling is then per-
formed in the decoder section to restore the original
size of the image and achieve high resolution segmen-
tation at the pixel level. Upsampling layers are used
here. In addition, there are skip connections between
the encoder and decoder to integrate the features of
the different layers of the encoder into the decoder.
The U-Net is a very lightweight network that has
already been used to segment trajectories in (Pohle-
Fr
¨
ohlich and Bolten, 2023). For the segmentation
with the U-Net, we used a network depth of four lay-
ers in combination with a loss function weighted by
the class frequency of the individual pixels.
4.4 Camera Calibration
To facilitate stereo vision, the camera heads must
first be calibrated. This serves the purposes of firstly
Stereo-Event-Camera-Technique for Insect Monitoring
379

(a) Points found in a single
detection.
(b) Average of multiple de-
tections.
Figure 9: Comparison of a single detection and averaged
object points.
undistorting the pixel coordinates, causing distances
between pixels to be unaffected by their positions in
the pixel array, and secondly determining the cam-
eras’ exact position relative to one another. This al-
lows us to align the event streams spatially and sim-
plify the matching of corresponding events later.
Typically, calibration is performed by detecting
readily discernible points of an object with a known
shape and determining the transformation matrices
matching these points’ detected pixel coordinates
with the presumed object point coordinates. For
stereo calibration, a transformation is computed that
maps the coordinate system of one camera to that of
the other in addition to the undistortion transforma-
tion.
Commonly, a chessboard pattern is used as the
known object, detecting the inside corners between
squares. For a frame camera, this can be a physical
chessboard of which photos are taken at various an-
gles and relative positions. Calibrating an event cam-
era introduces the complication that events are only
generated where there are changes in the scene. This
results in a static chessboard not being captured by
the camera. Moving a chessboard around in front
of the camera would introduce motion and make it
hard to acquire exactly corresponding object points.
To record a pattern with an event camera without
movement, it must change in brightness. One way to
achieve this is by displaying the calibration pattern on
a screen and making it flash. We displayed a flashing
black and white chessboard on a white background on
an IPS screen, causing events to be generated by the
black squares. The dimensions of the squares were
2.2 cm × 2.2 cm. We use the Metavision SDK im-
plementation (Prophesee, 2023a)
4
to detect the inside
corners in frames generated from 10 ms of events. To
mitigate inaccuracies in corner localization, we take
several static recordings of the chessboard from mul-
4
https://docs.prophesee.ai/stable/api/python/core ml/
corner detection.html
tiple angles and average the point positions of all de-
tections for each individual recording. The result of
this approach is illustrated in Figure 9.
Once the object points are acquired, intrinsic and
extrinsic camera parameters are calculated the same
way as they would be for a frame based camera. We
use the functionality provided by the Metavision SDK
to calculate each camera’s intrinsic parameters and
use OpenCV (Bradski, 2000) library functions to ac-
quire the extrinsic parameters based on the previously
acquired object points.
4.5 Depth Value Computation
The first step in the depth calculation after the cam-
era calibration is the rectification of the images to en-
sure that by reducing the correspondence problem by
the epipolar condition, the search for corresponding
points only has to be done along one image row. The
result is shown in Figure 10 for a part of the dataset.
The rectification is followed by a search for cor-
responding point pairs. For this purpose, the events
detected as insects in the two camera streams are pro-
jected into an image over a longer time period of em-
pirically determined 250 ms each, where the value at
the pixel position corresponds to the time stamp (see
(a) Plain display.
(b) Data after rectification.
Figure 10: Part of a dataset before and after the rectifica-
tion step (the frame indicates the undistorted section, and
the colors of the individual events correspond to the time
stamp).
VISAPP 2024 - 19th International Conference on Computer Vision Theory and Applications
380

(a) Image of the left camera.
(b) Image of the right camera.
(c) Image with the detected corresponding points.
Figure 11: Left and right images and detected correspond-
ing points.
Figure 11). The differences in the trajectories seen
in the figure between the left and right camera images
may be due to either differences in occlusion during
perspective projection from different viewing angles,
or differences in segmentation.
To determine depth, for each pixel in the right im-
age, a corresponding pixel in the left image is located,
including not only the same row but also two rows be-
low and above to account for calibration inaccuracies.
A pixel is considered corresponding if the time dif-
ference is minimal and less than 1.5 ms. This thresh-
old is empirically determined and compensates for in-
accuracies during synchronization. The difference of
the x-coordinate between the determined correspond-
ing values is stored as a disparity value. In the last
step, the 4D world coordinates (x, y, z, t) are calcu-
lated from these disparities and the reprojection ma-
trix determined during camera calibration, where the
coordinates are relative to the optical center of the left
camera.
5 RESULTS
To evaluate the different encoding methods with re-
spect to the quality of the segmentation, the dataset
described in Section 3.3 was used, which, however,
does not contain any stereo data. To evaluate the qual-
ity of the depth estimation, two additional datasets
were used. However, no segmentation ground truth
was available.
5.1 Segmentation Results
To investigate the segmentation quality, our dataset
of 12230 images was partitioned using 3690 images
as a test set. To ensure that the network had not
already seen immediately adjacent images, the total
number was first divided into blocks of 10 images
each, from which 369 blocks (30%) were randomly
selected. Each block contained a time interval of half
a second. Due to the fast movement of the insects, this
ensured sufficient variability between training and test
sets.
To compare segmentation results the F1 score was
used. The best results were obtained with histogram
encoding after 100 training epochs. The individual
results are shown in Table 2. The F1 scores for the
BACKGROUND class representing pixels in the en-
coding without any triggered events were 0.999 in all
cases and are therefore not included in the table.
When looking at the result images, it became clear
that the two classes of interest were mostly segmented
slightly too large, resulting in a low F1 value. There-
fore, only those class predictions where events ac-
tually occurred were considered in a post-processing
step. This makes sense because only these are impor-
tant for propagating the results back to the original
3D event stream for depth estimation (Pohle-Fr
¨
ohlich
and Bolten, 2023). The results of this post-processing
are given in Table 2b. The F1 values improved for all
classes and encodings. Again, the best results were
obtained for the histogram encoding.
Using the trained weights to predict events asso-
ciated with insect flight paths yields to similar re-
sults for two datasets taken from different views of
a meadow (Figure 13b) that were not included in the
training set. Figure 12 shows a section of the point
cloud visualizing all events for three of the used en-
coding methods. All points detected by event cube
encoding are colored red, all detected by optical flow
encoding are colored green, and all detected by his-
Stereo-Event-Camera-Technique for Insect Monitoring
381

Table 2: Resulting F1 scores for different encoding meth-
ods.
class Environ- Insect
ment
histogram 0.910 0.662
linear time surface 0.500 0.633
event cube 0.891 0.665
optical flow 0.490 0.663
(a) Plain inference after 100 epoch training.
class Environ- Insect
ment
histogram 0.967 0.897
linear time surface 0.956 0.857
event cube 0.912 0.755
optical flow 0.945 0.807
(b) Results after post-processing.
Figure 12: Result of the prediction for the dataset of the
meadow in direction 1 using the different encoding methods
(red: event cube encoding, green: optical flow encoding,
blue: histogram encoding). All other colors result from ad-
ditive color mixing. For the detected environmental events,
a value of 50 was additionally used for the alpha channel.
togram encoding are colored blue. All other colors
result from additive color mixing. Events shown in
white were predicted to be part of the insect flight path
by all three encodings. All black events were clas-
sified as noise or part of the plant movement by all
methods. It can be seen that the optical flow encod-
ing incorrectly predicted many events of plant move-
ment, while the event cube encoding often failed to
detect parts of the insect trajectory. The best results
(blue, white, magenta and cyan markers) are provided
by histogram encoding.
(a) All trajectories in 30 minutes for the meadow in view
direction 1.
(b) Considered meadow with the two directions of view
used.
Figure 13: Detected trajectories in a time interval of 30 min-
utes for the considered meadow shown below.
5.2 Depth Estimation Visualization
The visualization of the data from two 30-minute
recordings of a meadow in two different view direc-
tions is based on the computation of the 4D data us-
ing histogram encoding. Figure 13a shows all in-
sect movements over 30 minutes in the first record-
ing. In the displayed view volume, the time is coded
by the color. It can be seen that insect flight occurred
anywhere up to 4 meters in the meadow during the
recording.
Beyond about 4 m from the camera, the data
thinned out, partly due to inaccuracies in the camera
calibration and partly due to the chosen position of
the camera just below the tallest plants (Figure 13b).
The trajectories determined at shorter time intervals
for the two different views of the meadow are shown
in Figure 14 to better assess the quality of the results.
VISAPP 2024 - 19th International Conference on Computer Vision Theory and Applications
382

(a) 25 seconds of the first
recording.
(b) 90 seconds of the first
recording.
(c) 19 seconds of the second recording.
Figure 14: Trajectories of the first recording of a meadow
over 25 and 90 seconds and the second recording over 19
seconds.
6 CONCLUSIONS AND FUTURE
WORK
This paper presents the steps of the developed pro-
cessing pipeline from image acquisition to 4D display
for long-term insect monitoring with a stereo event
camera setup. Segmentation results have shown that
insect trajectories can be reliably separated from plant
movements. The use of histogram encoding gave the
best results. To improve the segmentation, besides
the improvement of the dataset a Siamese neural net-
work will be tested, which uses the same weights but
works in parallel on the two different input images to
obtain comparable segmentation results. This could
compensate for differences in segmentation quality
between the left and right camera images.
There are still some inaccuracies in the calcula-
tion of the 4D coordinates. These are caused by the
construction of the measurement system. The align-
ment of the two event cameras changed slightly due to
transport and heat, so that using the calibration data
resulted in an offset of up to 10 lines, depending on
the position of the events in the pixel matrix. A more
mechanically stable setup will be developed in the fu-
ture. For further interpretation of the data, the next
step will be to cluster the individual trajectories and
convert them to spline curves in order to obtain better
3D flight curves. Methods for trajectory tracking will
also be investigated in order to obtain longer sections
and to avoid double counting of insects. Finally, the
individual trajectories will be classified into different
insect groups based on the flight patterns as proposed
in (Pohle-Fr
¨
ohlich and Bolten, 2023).
REFERENCES
Bjerge, K., Mann, H. M., and Høye, T. T. (2022). Real-time
insect tracking and monitoring with computer vision
and deep learning. Remote Sensing in Ecology and
Conservation, 8(3):315–327.
Bolten, T., Lentzen, F., Pohle-Fr
¨
ohlich, R., and T
¨
onnies,
K. D. (2022a). Evaluation of deep learning based
3d-point-cloud processing techniques for semantic
segmentation of neuromorphic vision sensor event-
streams. In VISIGRAPP (4: VISAPP), pages 168–179.
Bolten, T., Pohle-Fr
¨
ohlich, R., and T
¨
onnies, K. (2023a). Se-
mantic Scene Filtering for Event Cameras in Long-
Term Outdoor Monitoring Scenarios. In Bebis, G.
et al., editors, 18th International Symposium on Vi-
sual Computing (ISVC), Advances in Visual Comput-
ing, volume 14362 of Lecture Notes in Computer Sci-
ence, pages 79–92, Cham. Springer Nature Switzer-
land.
Bolten, T., Pohle-Fr
¨
ohlich, R., and T
¨
onnies, K. D. (2023b).
Semantic segmentation on neuromorphic vision sen-
sor event-streams using pointnet++ and unet based
processing approaches. In VISIGRAPP (4: VISAPP),
pages 168–178.
Bolten, T., Pohle-Fr
¨
ohlich, R., Volker, D., Br
¨
uck, C.,
Beucker, N., and Hirsch, H.-G. (2022b). Visualiza-
tion of activity data from a sensor-based long-term
monitoring study at a playground. In VISIGRAPP (3:
IVAPP), pages 146–155.
Bradski, G. (2000). The OpenCV Library. Dr. Dobb’s Jour-
nal of Software Tools.
Dennis, R., Shreeve, T., Isaac, N., Roy, D., Hardy, P., Fox,
R., and Asher, J. (2006). The effects of visual ap-
parency on bias in butterfly recording and monitoring.
Biological conservation, 128(4):486–492.
Dong, S., Du, J., Jiao, L., Wang, F., Liu, K., Teng, Y.,
and Wang, R. (2022). Automatic crop pest detection
oriented multiscale feature fusion approach. Insects,
13(6):554.
Droissart, V., Azandi, L., Onguene, E. R., Savignac, M.,
Smith, T. B., and Deblauwe, V. (2021). Pict: A
low-cost, modular, open-source camera trap system to
study plant–insect interactions. Methods in Ecology
and Evolution, 12(8):1389–1396.
Gallego, G., Delbr
¨
uck, T., Orchard, G., Bartolozzi, C.,
Taba, B., Censi, A., Leutenegger, S., Davison, A. J.,
Conradt, J., Daniilidis, K., et al. (2020). Event-based
vision: A survey. IEEE transactions on pattern anal-
ysis and machine intelligence, 44(1):154–180.
Hallmann, C. A., Sorg, M., Jongejans, E., Siepel, H.,
Hofland, N., Schwan, H., Stenmans, W., M
¨
uller,
A., Sumser, H., H
¨
orren, T., et al. (2017). More
than 75 percent decline over 27 years in total fly-
ing insect biomass in protected areas. PloS one,
12(10):e0185809.
Stereo-Event-Camera-Technique for Insect Monitoring
383

Marcasan, L. I. S., HULUJAN, I. B., Florian, T., Somsai,
P. A., MILITARU, M., Sestras, A. F., Oltean, I., and
Sestras, R. E. (2022). The importance of assessing
the population structure and biology of psylla species
for pest monitoring and management in pear orchards.
Notulae Botanicae Horti Agrobotanici Cluj-Napoca,
50(4):13022–13022.
Pellegrino, N., Gharaee, Z., and Fieguth, P. (2022). Ma-
chine learning challenges of biological factors in in-
sect image data. arXiv preprint arXiv:2211.02537.
Pohle-Fr
¨
ohlich, R. and Bolten, T. (2023). Concept study
for dynamic vision sensor based insect monitoring.
In Radeva, P., Farinella, G. M., and Bouatouch, K.,
editors, Proceedings of the 18th International Joint
Conference on Computer Vision, Imaging and Com-
puter Graphics Theory and Applications, VISIGRAPP
2023, Volume 4: VISAPP, 2023, pages 411–418.
Prophesee (2023a). Documentation metavision sdk. https:
//docs.prophesee.ai/stable/index.html.
Prophesee (2023b). Metavision evaluation kit-3. https://
www.prophesee.ai/event-based-evk-3/.
Qing, Y., Jin, F., Jian, T., XU, W.-g., ZHU, X.-h., YANG,
B.-j., Jun, L., XIE, Y.-z., Bo, Y., WU, S.-z., et al.
(2020). Development of an automatic monitoring sys-
tem for rice light-trap pests based on machine vision.
Journal of Integrative Agriculture, 19(10):2500–2513.
Ratnayake, M. N., Dyer, A. G., and Dorin, A. (2021). To-
wards computer vision and deep learning facilitated
pollination monitoring for agriculture. In Proceedings
of the IEEE/CVF conference on computer vision and
pattern recognition, pages 2921–2930.
Ronneberger, O., Fischer, P., and Brox, T. (2015). U-Net:
Convolutional Networks for Biomedical Image Seg-
mentation. In International Conference on Medical
image computing and computer-assisted intervention,
pages 234–241. Springer.
Sutherland, W. J., Roy, D. B., and Amano, T. (2015). An
agenda for the future of biological recording for eco-
logical monitoring and citizen science. Biological
Journal of the Linnean Society, 115(3):779–784.
W
¨
agele, J. W., Bodesheim, P., Bourlat, S. J., Denzler,
J., Diepenbroek, M., Fonseca, V., Frommolt, K.-H.,
Geiger, M. F., Gemeinholzer, B., Gl
¨
ockner, F. O., et al.
(2022). Towards a multisensor station for automated
biodiversity monitoring. Basic and Applied Ecology,
59:105–138.
VISAPP 2024 - 19th International Conference on Computer Vision Theory and Applications
384
