
Triple-stream Deep Metric Learning of Great Ape Behavioural Actions
Otto Brookes
1
, Majid Mirmehdi
1
, Hjalmar K
¨
uhl
2
and Tilo Burghardt
1
1
Department of Computer Science, University of Bristol, U.K.
2
Evolutionary and Anthropocene Ecology, iDiv, Leipzig, Germany
tilo@cs.bris.ac.uk, hjalmar.kuehl@idiv.de
Keywords:
Animal Biometrics, Multi-Stream Deep Metric Learning, Animal Behaviour, Great Apes, PanAf-500 Dataset.
Abstract:
We propose the first metric learning system for the recognition of great ape behavioural actions. Our proposed
triple stream embedding architecture works on camera trap videos taken directly in the wild and demonstrates
that the utilisation of an explicit DensePose-C chimpanzee body part segmentation stream effectively com-
plements traditional RGB appearance and optical flow streams. We evaluate system variants with different
feature fusion techniques and long-tail recognition approaches. Results and ablations show performance im-
provements of ∼ 12% in top-1 accuracy over previous results achieved on the PanAf-500 dataset containing
180,000 manually annotated frames across nine behavioural actions. Furthermore, we provide a qualitative
analysis of our findings and augment the metric learning system with long-tail recognition techniques showing
that average per class accuracy – critical in the domain – can be improved by ∼ 23% compared to the literature
on that dataset. Finally, since our embedding spaces are constructed as metric, we provide first data-driven
visualisations of the great ape behavioural action spaces revealing emerging geometry and topology. We hope
that the work sparks further interest in this vital application area of computer vision for the benefit of endan-
gered great apes. We provide all key source code and network weights alongside this publication.
1 INTRODUCTION
As the climate crisis gathers pace, the threat to many
endangered species grows ever more perilous (Al-
mond et al., 2022). All species of great apes are, for
instance, listed as endangered or critically endangered
according to the IUCN Red List (IUCN, 2022).
Consequently, there is urgent need for methods
that can help to monitor population status and assess
the effectiveness of conservation interventions (K
¨
uhl
and Burghardt, 2013; Congdon et al., 2022; Tuia
et al., 2022). This includes the recognition of behav-
iors and variation therein, as an integral part of biolog-
ical diversity (Dominoni et al., 2020; Carvalho et al.,
2022).
Previous works have employed deep neural net-
works which leverage multiple modalities, such as
RGB, optical flow, and audio (Sakib and Burghardt,
2020; Bain et al., 2021), for the classification of great
ape behaviours and actions. However, higher level ab-
stractions such as pose or body part information have
remained unexplored for addressing this task. In re-
sponse, we propose utilising the latter together with
RGB and optical flow in a triple-stream metric learn-
ing system (see Fig. 1) for improved classification re-
sults and domain visualisations relevant to biologists.
Great Ape Activities. This paper will focus on
great ape activity recognition, where the coarse ac-
tivity classes used are illustrated in Fig. 2 for the
utilised PanAf-500 dataset (see Sec. 3). Note that
computer vision would traditionally categorise these
classes as actions whilst in the biological realm they
represent behaviour (or aspects thereof) often cap-
tured in ethograms (Nishida et al., 1999; Zamma and
Matsusaka, 2015). For clarity, in this paper we will
refer to these classes as behavioural actions recognis-
ing historical traditions in both disciplines.
We will approach the classification task via a deep
metric learning system (Karaderi et al., 2022) that
embeds inputs into a latent space and uses geomet-
ric distances to form distributions that align with the
semantic similarity captured by the classes (Hermans
et al., 2017; Musgrave et al., 2020). A major advan-
tage over standard supervised systems is that sample
distances in visualisations of the latent space always
relate to learned similarity and, thus, are more natu-
rally interpretable by experts. We will also analyse
294
Brookes, O., Mirmehdi, M., Kühl, H. and Burghardt, T.
Triple-stream Deep Metric Learning of Great Ape Behavioural Actions.
DOI: 10.5220/0011798400003417
In Proceedings of the 18th International Joint Conference on Computer Vision, Imaging and Computer Graphics Theory and Applications (VISIGRAPP 2023) - Volume 5: VISAPP, pages
294-302
ISBN: 978-989-758-634-7; ISSN: 2184-4321
Copyright
c
2023 by SCITEPRESS – Science and Technology Publications, Lda. Under CC license (CC BY-NC-ND 4.0)
positive
anchor
fusion
negative
ResNet-50
ResNet-50
ResNet-50
x
0
.
.
.
.
.
.
x
128
x
0
.
.
.
.
.
.
x
128
x
0
.
.
.
.
.
.
x
128
Metric
Learning
d
d
d
d
L
Triplet
x
0
.
.
.
.
.
.
x
128
x
0
.
.
.
.
.
.
x
128
embedding model
DensePose-C
Optical flow
RGB
x
0
.
.
.
.
.
.
x
128
x
0
.
.
.
.
.
.
x
128
x
0
.
.
.
.
.
.
x
128
x
0
.
.
.
.
.
.
x
128
x
0
.
.
.
.
.
.
x
128
x
0
.
.
.
.
.
.
x
128
x
0
.
.
.
.
.
.
x
128
embedding model
embedding model
shared weights
shared weights
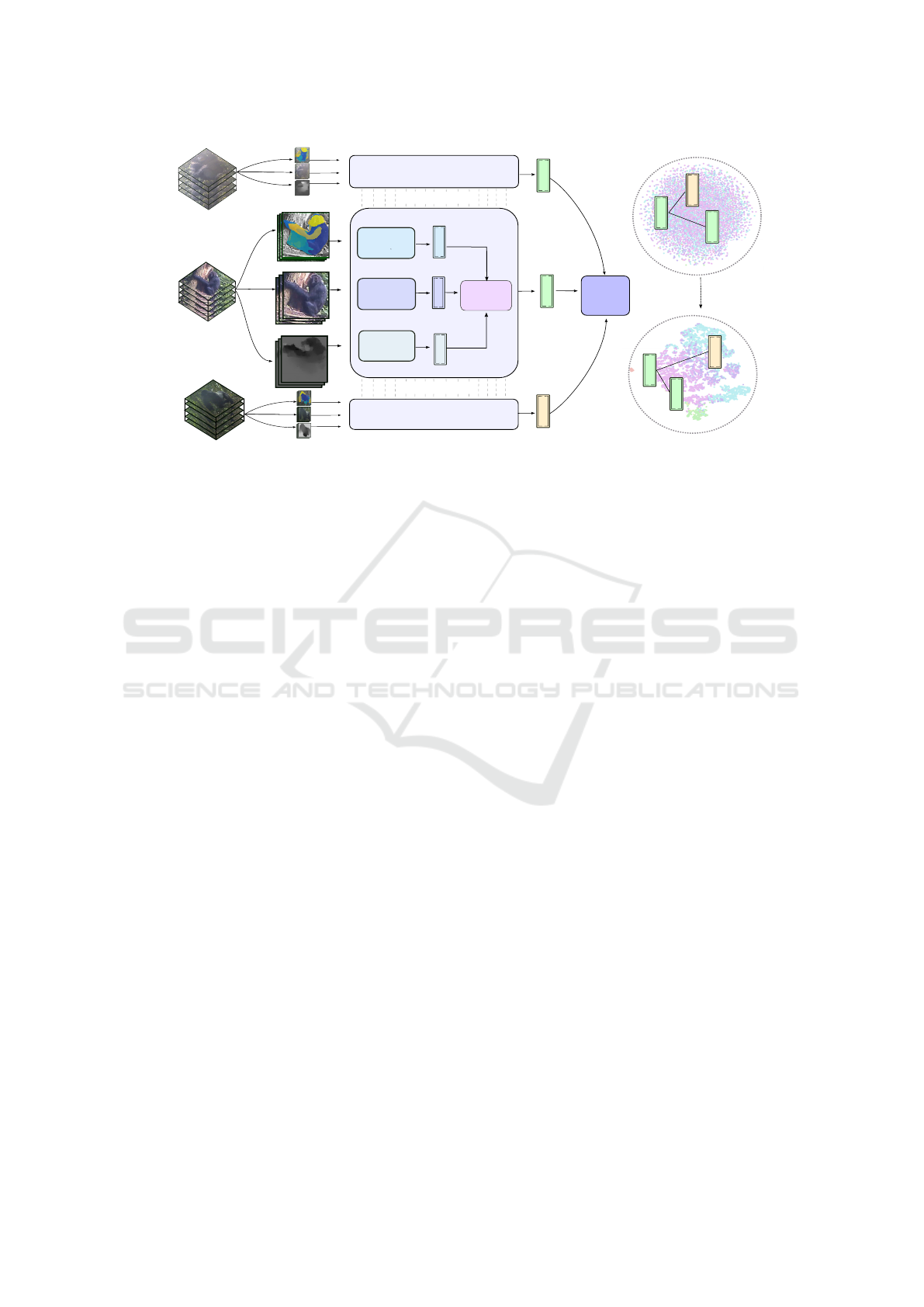
Figure 1: System Overview. Our proposed triple-stream metric learning approach utilises all RGB appearance, optical flow,
and DensePose-C segmentations of chimps in videos. Exploiting hybrid reciprocal triplet and cross entropy losses, the model
is then trained to map embeddings representing great ape behavioural actions onto a metric space, where semantically similar
representations are geometrically close forming natural clusters. This pipeline improves on state-of-the-art classification
performance and allows for visualisations of the underpinning space of behavioural actions (best viewed zoomed).
the role that additional DensePose-Chimp informa-
tion (Sanakoyeu et al., 2020) can play in improving
recognition performance compared to systems that
utilise RGB and optical flow only. Lastly, as shown
by Sakib and Burghardt (Sakib and Burghardt, 2020),
there are significant challenges in correctly classify-
ing behavioural actions which occur infrequently and
form the distribution tail (see Fig. 2). To address this,
we will employ three long-tailed recognition (LTR)
techniques to improve performance on tail classes; (i)
logit adjustment (Menon et al., 2020); (ii) class bal-
anced focal loss (Cui et al., 2019); and (iii) weight
balancing (Alshammari et al., 2022).
In summary, our contributions are as follows:
(i) we implement the first deep metric learning system
for recognising great ape behavioural actions; (ii) we
show that utilising explicit pose information has a sig-
nificant positive effect on recognition performance in
this domain; and (iii) we establish that existing LTR
techniques can be applied in a metric learning setting
to improve performance on tail classes for the prob-
lem. The proposed approaches improve the state-of-
the-art performance benchmarks with respect to top-1
(∼ 85%) and average per class (∼ 65%) accuracy on
the PanAf-500 dataset.
2 RELATED WORK
Action recognition aims to classify actions observed
in video (Kalfaoglu et al., 2020; Shaikh and Chai,
2021). Learning spatio-temporal features character-
istic for actions (Simonyan and Zisserman, 2014) via
various deep learning paradigms forms the approach
of choice in the domain of human action recogni-
tion (HAR). We will briefly review concepts from this
field, before discussing specifc relevant great ape be-
havioural action recognition and LTR methods.
Human Action Recognition. Although there are
numerous deep learning approaches to action recog-
nition (Zhou et al., 2018; Lin et al., 2019; Tran et al.,
2019; Kalfaoglu et al., 2020; Pan et al., 2019; Majd
and Safabakhsh, 2020; Sharir et al., 2021; Zhang
et al., 2021a) this work focuses on multi-stream ar-
chitectures, which address key aspects of the action
recognition problem (e.g., spatial and temporal) in-
dependently and explicitly. Feichtenhofer et al. (Fe-
ichtenhofer et al., 2019) introduced the SlowFast ar-
chitecture which employs two streams, each operat-
ing at different frame rates; a slow, low frame-rate
pathway captures spatial information while the fast,
high frame-rate pathway captures fine temporal detail.
Other types of multi-stream networks process differ-
ent visual modalities. Simonyan (Simonyan and Zis-
serman, 2014) introduced a two-stream network that
processes RGB and optical flow to exploit spatial and
temporal semantics, respectively. Since then, several
networks that utilise additional modalities, such as
motion saliency (Zong et al., 2021) and audio (Wang
et al., 2021), have been introduced. Recently, the in-
troduction of pose, which is critical for the perception
of actions (Le et al., 2022), has shown promising re-
sults in multi-stream architectures (Hong et al., 2019;
Triple-stream Deep Metric Learning of Great Ape Behavioural Actions
295

10
4
10
5
# Samples (log)
Behavioural action classes
hanging
walking
sitting on back
standing
sitting
climbing up
camera interaction
running
climbing down
Figure 2: Behavioural Actions in the PanAf-500 Data.
Examples of each one of the nine behavioural action classes
(top) and their distribution across the approx. 180k frames
in the dataset (bottom). Note the imbalance of two orders of
magnitude in the distribution (best viewed zoomed).
Hayakawa and Dariush, 2020; Duan et al., 2021; Li
et al., 2022). In particular, the DensePose format
provides an opportunity to exploit fine-grained, seg-
mentation map-based pose representations for action
recognition. Hayakawa et al. (Hayakawa and Dar-
iush, 2020) combine RGB and DensePose estimations
in a two-stream network and demonstrate strong per-
formance on egocentric footage of humans. Whilst
such significant progress has been made in the domain
of HAR, research into great ape behavioural action
recognition is still in its infancy and few systems have
been tested on natural datasets.
Great Ape Domain. To date, two systems have
attempted automated great ape behavioural action
recognition, both are multi-stream architectures. The
first (Sakib and Burghardt, 2020) is based on the
two-stream convolutional architecture by Simonyan
et al. (Simonyan and Zisserman, 2014) and used 3D
ResNet-18s for feature extraction and LSTM-based
fusion of RGB and optical flow features. They report
top-1 accuracy of 73.52% across the nine behavioural
actions in the PanAf-500 dataset (see Sec. 3) and a
relatively low average per class accuracy (42.33%),
highlighting the issue of tail class performance. The
second, proposed by Bain et al. (Bain et al., 2021),
is a deep learning system that requires both audio
and video inputs and detects two specific behaviours;
buttress drumming and nut cracking. Their system
utilised a 3D ResNet-18 and a 2D ResNet-18 for ex-
traction of visual and assisting audio features, respec-
tively, in different streams. They achieved an aver-
age precision of 87% for buttress drumming and 85%
for nut cracking on their unpublished dataset. How-
ever, the multi-modal method is not applicable to all
camera trap settings since many older models do not
provide audio. It cannot be utilised on the PanAf-500
dataset since many clips there do not contain audio.
Long-Tailed Recognition. Most natural recorded
data exhibits long-tailed class distributions (Liu et al.,
2019). This is true of great ape camera-trap footage
which is dominated by commonly occurring be-
haviours - even with only the nine classes of the
PanAf-500 data the distribution shows a clear tail (see
Fig. 2). Without addressing this issue, models trained
on such data often exhibit poor performance on rare
classes. Various counter-measures have been pro-
posed (Verma et al., 2018; Kang et al., 2019; Zhang
et al., 2021b). Class balanced losses assign addi-
tional weights, typically determined by inverse class
frequencies, to samples from rare classes and have
yielded strong results when coupled with techniques
to reduce per-class redundancy (Cui et al., 2019).
Similarly, logit adjustment uses class frequencies to
directly offset output logits in favour of minority
classes during training (Menon et al., 2020). An
orthogonal approach, based on the observation that
weight norms for rare classes are smaller in naively
trained classifiers, is to perform weight balancing (Al-
shammari et al., 2022). These techniques have
achieved strong results on several LTR benchmarks.
Before detailing how we use triple-stream metric
learning with explicit DensePose-Chimp processing
and LTR extensions for behavioural action recogni-
tion, we will briefly outline the utilised dataset.
3 DATASET
The Pan-African dataset, gathered by the Pan African
Programme: ‘The Cultured Chimpanzee’, comprises
∼ 20,000 videos from footage gathered at 39 study
sites spanning 15 African countries. Here we utilise
a 500 video subset, PanAf-500, specifically ground-
truth labelled for use in computer vision under re-
VISAPP 2023 - 18th International Conference on Computer Vision Theory and Applications
296

Figure 3: Frame-by-frame Ground Truth Annotations.
Four still frames from PanAf-500 videos with annotations
of location (green boxes) and behavioural actions (visu-
alised as text) of the apes in-frame (best viewed zoomed).
producible and comparable benchmarks. It includes
frame-by-frame annotations for full-body locations of
great apes and nine behavioural actions (Sakib and
Burghardt, 2020) across approximately 180k frames
(see. Fig. 3). Fig. 2 displays the behavioural actions
classes in focus together with their distribution. We
utilised the PanAf-500 dataset for all experiments and
employ the same training and test partitions described
in (Sakib and Burghardt, 2020).
4 METHOD
The proposed system utilises three visual modali-
ties as input; RGB, optical flow, and DensePose-C
estimations (Sanakoyeu et al., 2020), as illustrated
in Fig. 1). All optical flow images are pre-computed
using OpenCV’s implementation of the Dual TV L1
algorithm (Zach et al., 2007). We employ the model
developed by Sanakoyeu et al. (Sanakoyeu et al.,
2020) to generate DensePose-C segmentations de-
scribing chimpanzee pose. The model predicts dense
correspondences between image pixels and a 3-D ob-
ject mesh where each mesh represents a chimpanzee
body part specified by a selector I and local surface
coordinates within each mesh indexed by U and V .
Frame-by-frame application to each of the PanAf-
500 videos yields DensePose-C estimates expressed
in IUV coordinates.
Each of the three input modalities is fed into a 3D
ResNet-50 (Du Tran et al., 2017) backbone, which
together act as a feature extractor (see Fig. 1). The
input tensors into the backbones are 3D since inputs
are processed in snippets, that is each stream accepts a
sequence of n consecutive RGB frames, optical flow
images, or IUV coordinates, respectively. The final
fully-connected layer outputs an n-dimensional en-
coding for each stream. These are fused into a single
embedding using three popular approaches; (i) sim-
x
0
.... x
128
Concatenation
Conv3D
ResNet50
ResNet50
ResNet50
b x 6144 x 5 x 8 x 8
MaxPool3D
Feature maps
2048 x 5 x 8 x 8
feature extractors
Triple stream
AdaptiveAvgPool3D
Fc1
b x 2048 x 3 x 5 x 5
Fc2
b x 1024
x
0
.... x
128
x
0
.... x
128
x
0
.... x
128
Multiplication
L2 norm
x
0
.... x
128
RGB
Optical flow
Output
DensePose-C
Figure 4: Fusion Head Schematics. A component break-
down of fusion by element-wise multiplication (left) and
convolutional fusion (right) as applied for our work to ex-
plore their impact on performance.
ple averaging across streams; (ii) convolutional fusion
whereby stream features are concatenated and passed
to a 3D convolutional layer as a volume; and (iii)
element-wise multiplication of all three embedding
vectors followed by L2 normalisation. The latter two
approaches are illustrated in detail in Fig. 4. A lin-
ear layer at the end of the fusion head finally outputs
the unified embedding as logits. Whilst this system
was trained via metric learning - visually sketched in
Fig. 1 (right) - a k-NN classifier is used to perform
inference in the embedding space during evaluation.
Let the parameters of this network f
θ
(·) be de-
noted by θ. Furthermore, let f
θ
(x) = x be the short-
hand for referring to embeddings. Our metric learn-
ing objective is, thus, to minimise the distance be-
tween anchor-positive embedding pairs d(x
a
, x
p
) and
maximise distance between anchor-negative embed-
ding pairs d(x
a
, x
n
), where d represents a Euclidean.
Instead of using standard triplet loss (Hermans et al.,
2017) L
T L
, we use an improved version (Andrew
et al., 2021), where the model is optimised via a hy-
brid reciprocal triplet and softmax cross-entropy loss:
L
RC
= L
CE
+ λ L
RT
. (1)
It is assembled from two components balanced by
λ = 0.1 as given in (Andrew et al., 2021). The two
Triple-stream Deep Metric Learning of Great Ape Behavioural Actions
297

components themselves are evaluated as:
L
RT
= d(x
a
, x
p
) +
1
d(x
a
, x
n
)
(2)
L
CE
= −log
e
x
y
∑
C
i=1
e
x
i
, (3)
where C denotes the total number of classes and y are
the class labels.
In order to extend this system into the LTR do-
main we substitute the softmax cross-entropy term
for losses calculated using; (i) cross-entropy soft-
max with logit adjustment (Menon et al., 2020) L
LA
;
(ii) class-balanced focal loss (Cui et al., 2019) L
CB
;
and (iii) class-balanced focal loss with weight balanc-
ing (Alshammari et al., 2022). The first two losses are
evaluated as follows:
L
LA
= −log
e
x
y
+ τ · log π
y
∑
C
i=1
e
x
i
+τ · log π
i
, (4)
L
CB
= −
1 − β
1 − β
n
y
C
∑
i=1
(1 − p
i
)
γ
log(p
i
), (5)
where π represents the class priors (i.e., class frequen-
cies in the training set) and temperature factor τ = 1,
β = 0.99 is the re-weighting hyper-parameter, n is the
total number of samples, y are the classes, γ = 1 is the
focal loss hyper-parameter and p
i
= σ(x
i
). Balancing
the network weights θ is performed via a MaxNorm
constraint ∥θ
l,i
∥
2
2
≤ δ
2
, ∀i given in (Alshammari et al.,
2022) imposed on each class filter i in the last layer l
of the network where δ is the L2-norm ball radius. We
will reference a L
CB
-based optimisation where weight
balancing is performed via L
W B
.
Methodologically, this described architecture ap-
proaches the learning of behavioural great ape actions
via five key capabilities: 1) utilisation of multiple rel-
evant input modalities across an entire video snippet;
2) effective streamed content encoding; 3) fusion into
a single embedding space; 4) metric space optimisa-
tion so that distances naturally reflect semantic sim-
ilarity; and 5) taking into account class imbalances
common to the domain content.
5 EXPERIMENTS
5.1 General Training Setup
We train our architecture via SGD optimisation using
batch size 32 and learning rate 10
−4
. Feature extrac-
tor backbones are initialised with Kinetics-400 (Kay
et al., 2017) pre-trained weights and training runs are
distributed over 8 Tesla V100 GPUs for 100 epochs.
Table 1: Behavioural Action Recognition Benchmarks.
Top-1 and average per-class (C-Avg) accuracy performance
on the PanAf-500 dataset for the current state-of-the-
art (row 1), single and dual-stream baselines (rows 2–5),
and our triple-stream networks (rows 6–8) for different fu-
sion methodologies and losses tested.
Models/Streams Fusion Loss Top-1 C-Avg
Sakib et al. 2020
1 RGB+OF LSTM L
FL
73.52% 42.33%
Up to Dual-Stream
2 RGB only None L
T L
55.50% 32.67%
3 RGB only None L
RC
74.24% 55.76%
4 RGB+OF Avg L
T L
62.90% 39.10%
5 RGB+OF Avg L
RC
75.02% 61.97%
Triple-Stream (Ours)
6 RGB+OF+DP Avg L
RC
81.71% 46.61%
7 RGB+OF+DP Conv L
RC
82.04% 56.31%
8 RGB+OF+DP Elem L
RC
85.86% 50.50%
5.2 Baselines and Stream Ablations
As shown in Tab. 1, we first establish performance
benchmarks for one and two stream baseline archi-
tectures of our system (rows 2–5) against the cur-
rent state-of-the-art (row 1), which uses a ResNet-18
backbone with focal loss L
FL
, SGD, and LSTM-based
frame fusion (Sakib and Burghardt, 2020). As ex-
pected, we confirmed that - using identical setups and
losses - adding an optical flow stream is beneficial
in the great ape domain mirroring HAR results (see
rows 2 vs 4, and 3 vs 5). Additionally, models trained
using L
RC
consistently outperformed standard triplet
loss L
RC
scenarios (see rows 2 vs 3, and 4 vs 5). Fi-
nally, a dual-stream version of our proposed architec-
ture trained with L
RC
outperforms the state-of-the-art
by a small margin (see rows 1 vs 5).
5.3 Triple-Stream Recognition
As given in Tab. 1 rows 6–8, our proposed triple-
stream architecture significantly outperforms all base-
lines with regards to top-1 accuracy, achieving up to
85.86%. Thus, explicit DensePose-C information ap-
pears a useful information source for boosting be-
havioural action recognition in great apes. However,
without LTR techniques all our triple-stream models
are significantly outperformed by a dual-stream set-
ting (row 5) with regards to average per-class accu-
racy. This reduction is caused by significantly poorer
performance on minority classes (see Sec. 5.4).
Since the learned behavioural action embeddings
are constructed as metric from the outset, they can
be visualised meaningfully – we note that such data-
driven visualisations are novel in the primatology do-
VISAPP 2023 - 18th International Conference on Computer Vision Theory and Applications
298

camera interaction
climbing up
climbing down
hanging
running
sitting
sitting on back
standing
walking
Behavioural actions
Single Stream (RGB)
Kinetics pretrained (no training)
Training
Training
Test
Dual Stream (RGB+OF)
Triple Stream (AllThree)
Figure 5: Visualisations of Great Ape Behavioural Action Spaces. A 2D t-SNE (Wattenberg et al., 2016) visualisation of
the 128-dimensional training (top-right) and test (bottom-right) embeddings produced by the single, dual and three-stream
network with convolutional fusion. We can see that training set embeddings from all classes are clustered cleanly. In contrast,
test set embeddings show significant overlap and only embeddings from majority classes form distinct clusters. This is
consistent with the high top-1 accuracy and relatively low average per-class accuracy reported in Tab. 1.
main. Fig. 5 depicts such learned spaces for our data
and architecture where, independent of stream cardi-
nality, embeddings cluster the training data cleanly.
This is of course expected given above 99% top-1
training accuracy in all settings. Yet, behavioural ac-
tions of great apes are highly intricate as well as vari-
able and, even with approx. 144, 000 training frames
used, the model clearly shows signs of overfitting. As
a result, test set embeddings exhibit significant cluster
overlap. Sample groups representing sitting, standing,
and walking, for instance, blend into one another. In
addition to overfitting, this also highlights the transi-
tional nature of these often temporarily adjacent and
smoothly changing actions. Thus, future temporally
transitional ground truth labelling may be needed to
represent behavioural great ape action in the PanAf-
500 dataset more authentically.
5.4 Fusing Streams
When looking at the impact of information fusion
methods on performance in more detail, we find that
benchmarks vary significantly (see Tab. 1 rows 6–8)
when we test averaging, element-wise multiplication,
and convolutional fusion, as described in Sec. 4. Re-
sults show that convolution and element-wise mul-
tiplication improve performance slightly across both
metrics when compared with averaging: top-1 accu-
racy improves by 0.33% and 4.1%, respectively (see
rows 6–8). However, the most significant gains are
observed with respect to average per class accuracy
which increases by 3.44% for element-wise multipli-
cation and 9.7% for convolutional fusion. Learnable
parameters in the convolution method clearly help
blending information even when only fewer samples
are available for training. Building on this improve-
ment, we will next investigate the impact of LTR
methods in order to benefit tail class performance.
5.5 Long-Tail Recognition
When grouping behavioural actions into head (cov-
ering sitting, standing, and walking) and remain-
ing tail classes based on frequency in the data (see
Fig. 2), a significant performance gap becomes appar-
ent even when using the so far best C-Avg performing
model (see Tab. 2 row 1). Employing LTR techniques
can, however, reduce this gap and improve average
per-class accuracy further as quantified across rows
2–4 in Tab. 2). Fig. 6 shows t-SNE visualisations of
the three LTR triple-stream approaches when trained
with convolutional feature fusion. Particularly for the
class-balanced approaches and weight-balancing se-
tups (two rightmost), tail class clusters appear more
clearly separated and class overlap is generally re-
duced. Thus, for the great ape domain underrepre-
sented classes are indeed an effective source of infor-
mation for improving action separability in general.
Triple-stream Deep Metric Learning of Great Ape Behavioural Actions
299

climbing up
hanging
running
sitting
sitting on back
standing
walking
Logit adjustment
No LTR augmentation
Weight balanced
CB (+focal loss)
climbing down
camera interaction
Test
Figure 6: Long-tail Test Embeddings. A 2D t-SNE visualisation of the 128-dimensional test embeddings produced by the
three-stream network with convolutional fusion alone (leftmost) and augmented with each LTR technique; (i) logit adjustment
(ii) CB (+focal loss) and (iii) weight balancing. All LTR-augmented methods improve clustering of embeddings belonging to
tail classes. They appear more clearly separated and exhibit less overlap when compared with the non-LTR method.
6 CONCLUSION
In this work we introduced the first deep metric learn-
ing system for great ape behavioural action recogni-
tion. We demonstrated that the proposed triple-stream
architecture can provide leading state-of-the-art per-
formance when tested on the PanAf-500 camera trap
dataset covering 180,000 annotated frames across 500
videos taken in the wild. We demonstrated that the ad-
dition of a DensePose-C chimpanzee pose estimation
stream into the embedding architecture is highly ef-
fective and leads to system performance of 85.86%
top-1 accuracy on the data. We also showed that
adding LTR techniques that address poor tail class
performance to the system can improve the average
per-class accuracy to 65.66% on the dataset. Despite
these improvements we note that both larger anno-
tated datasets to counteract overfitting as well as more
temporally blended forms of annotation (e.g. action
transition annotations) would benefit the authenticity
of data-driven great ape behavioural representations.
We hope that the research presented here sparks fur-
ther interest in this vital application area for the bene-
fit of endangered species such as great apes.
ACKNOWLEDGEMENTS
We thank the Pan African Programme: ‘The Cul-
tured Chimpanzee’ team and its collaborators for al-
lowing the use of their data for this paper. We thank
Amelie Pettrich, Antonio Buzharevski, Eva Martinez
Garcia, Ivana Kirchmair, Sebastian Sch
¨
utte, Linda
Gerlach and Fabina Haas. We also thank manage-
ment and support staff across all sites; specifically
Yasmin Moebius, Geoffrey Muhanguzi, Martha Rob-
bins, Henk Eshuis, Sergio Marrocoli and John Hart.
Thanks to the team at https://www.chimpandsee.org
particularly Briana Harder, Anja Landsmann, Laura
K. Lynn, Zuzana Mach
´
a
ˇ
ckov
´
a, Heidi Pfund, Kris-
teena Sigler and Jane Widness. The work that al-
lowed for the collection of the dataset was funded
by the Max Planck Society, Max Planck Society In-
novation Fund, and Heinz L. Krekeler. In this re-
spect we would like to thank: Ministre des Eaux et
For
ˆ
ets, Minist
`
ere de l’Enseignement sup
´
erieur et de la
Recherche scientifique in C
ˆ
ote d’Ivoire; Institut Con-
golais pour la Conservation de la Nature, Minist
`
ere de
la Recherche Scientifique in Democratic Republic of
Congo; Forestry Development Authority in Liberia;
Direction Des Eaux Et For
ˆ
ets, Chasses Et Conserva-
tion Des Sols in Senegal; Makerere University Bi-
ological Field Station, Uganda National Council for
Science and Technology, Uganda Wildlife Authority,
National Forestry Authority in Uganda; National In-
stitute for Forestry Development and Protected Area
Management, Ministry of Agriculture and Forests,
Ministry of Fisheries and Environment in Equatorial
Guinea. This work was supported by the UKRI CDT
in Interactive AI under grant EP/S022937/1.
Table 2: LTR-enabled Behavioural Action Recogni-
tion Benchmarks. Average per-class accuracy for our
triple-stream network with convolutional fusion for best
performing non-LTR method (row1), and three LTR ap-
proaches (rows 2–4) targetting poor tail class performance.
Method/Loss C-Avg Head Tail
Non-LTR Triple-Stream
1 L
RC
56.31 80.57 44.78
LTR Triple-Stream
2 L
LA
61.76 83.22 50.7
3 L
CB
63.56 77.60 55.95
4 L
W B
65.66 82.55 56.26
VISAPP 2023 - 18th International Conference on Computer Vision Theory and Applications
300

REFERENCES
Almond, R., Grooten, M., Juffe Bignoli, D., and Petersen,
T. (2022). Wwf (2022) living planet report 2022 -
building a nature-positive society. 1
Alshammari, S., Wang, Y.-X., Ramanan, D., and Kong, S.
(2022). Long-tailed recognition via weight balancing.
In CVPR, pages 6897–6907. 2, 3, 5
Andrew, W., Gao, J., Mullan, S., Campbell, N., Dowsey,
A. W., and Burghardt, T. (2021). Visual identification
of individual holstein-friesian cattle via deep metric
learning. Computers and Electronics in Agriculture,
185:106133. 4
Bain, M., Nagrani, A., Schofield, D., Berdugo, S., Bessa, J.,
Owen, J., Hockings, K. J., Matsuzawa, T., Hayashi,
M., Biro, D., et al. (2021). Automated audiovisual
behavior recognition in wild primates. Science ad-
vances, 7(46):eabi4883. 1, 3
Carvalho, S., Wessling, E. G., Abwe, E. E., Almeida-
Warren, K., Arandjelovic, M., Boesch, C., Danquah,
E., Diallo, M. S., Hobaiter, C., Hockings, K., et al.
(2022). Using nonhuman culture in conservation re-
quires careful and concerted action. Conservation Let-
ters, 15(2):e12860. 1
Congdon, J., Hosseini, M., Gading, E., Masousi, M.,
Franke, M., and MacDonald, S. (2022). The future
of artificial intelligence in monitoring animal identifi-
cation, health, and behaviour. 1
Cui, Y., Jia, M., Lin, T.-Y., Song, Y., and Belongie, S.
(2019). Class-balanced loss based on effective num-
ber of samples. In CVPR, pages 9268–9277. 2, 3,
5
Dominoni, D. M., Halfwerk, W., Baird, E., Buxton, R. T.,
Fern
´
andez-Juricic, E., Fristrup, K. M., McKenna,
M. F., Mennitt, D. J., Perkin, E. K., Seymoure, B. M.,
et al. (2020). Why conservation biology can benefit
from sensory ecology. Nature Ecology & Evolution,
4(4):502–511. 1
Du Tran, H. W., Torresani, L., Ray, J., Lecun, Y., and Paluri,
M. (2017). A closer look at spatiotemporal convolu-
tions for action recognition.(2017). OK. 4
Duan, M., Qiu, H., Zhang, Z., and Wu, Y. (2021). Ntu-
densepose: A new benchmark for dense pose action
recognition. In Big Data, pages 3170–3175. IEEE. 3
Feichtenhofer, C., Fan, H., Malik, J., and He, K. (2019).
Slowfast networks for video recognition. In ICCV,
pages 6202–6211. 2
Hayakawa, J. and Dariush, B. (2020). Recognition and
3d localization of pedestrian actions from monocular
video. In ITSC, pages 1–7. IEEE. 3
Hermans, A., Beyer, L., and Leibe, B. (2017). In defense
of the triplet loss for person re-identification. arXiv
preprint arXiv:1703.07737. 1, 4
Hong, J., Cho, B., Hong, Y. W., and Byun, H. (2019).
Contextual action cues from camera sensor for multi-
stream action recognition. Sensors, 19(6):1382. 3
IUCN (2022). Iucn red list of threatened species version
2022.1. 1
Kalfaoglu, M. E., Kalkan, S., and Alatan, A. A. (2020).
Late temporal modeling in 3d cnn architectures with
bert for action recognition. In ECCV, pages 731–747.
Springer. 2
Kang, B., Xie, S., Rohrbach, M., Yan, Z., Gordo, A., Feng,
J., and Kalantidis, Y. (2019). Decoupling representa-
tion and classifier for long-tailed recognition. arXiv
preprint arXiv:1910.09217. 3
Karaderi, T., Burghardt, T., Hsiang, A. Y., Ramaer, J., and
Schmidt, D. N. (2022). Visual microfossil identifica-
tion via deep metric learning. In ICPRAI, pages 34–
46. Springer. 1
Kay, W., Carreira, J., Simonyan, K., Zhang, B., Hillier, C.,
Vijayanarasimhan, S., Viola, F., Green, T., Back, T.,
Natsev, P., et al. (2017). The kinetics human action
video dataset. arXiv preprint arXiv:1705.06950. 5
K
¨
uhl, H. S. and Burghardt, T. (2013). Animal biomet-
rics: quantifying and detecting phenotypic appear-
ance. TREE, 28(7):432–441. 1
Le, V.-T., Tran-Trung, K., and Hoang, V. T. (2022). A com-
prehensive review of recent deep learning techniques
for human activity recognition. Computational Intel-
ligence and Neuroscience, 2022. 2
Li, Y., Lu, Z., Xiong, X., and Huang, J. (2022). Perf-net:
Pose empowered rgb-flow net. In WACV, pages 513–
522. 3
Lin, J., Gan, C., and Han, S. (2019). Tsm: Temporal shift
module for efficient video understanding. In ICCV,
pages 7083–7093. 2
Liu, Z., Miao, Z., Zhan, X., Wang, J., Gong, B., and Yu,
S. X. (2019). Large-scale long-tailed recognition in
an open world. In CVPR, pages 2537–2546. 3
Majd, M. and Safabakhsh, R. (2020). Correlational con-
volutional lstm for human action recognition. Neuro-
computing, 396:224–229. 2
Menon, A. K., Jayasumana, S., Rawat, A. S., Jain, H., Veit,
A., and Kumar, S. (2020). Long-tail learning via logit
adjustment. arXiv preprint arXiv:2007.07314. 2, 3, 5
Musgrave, K., Belongie, S., and Lim, S.-N. (2020). Pytorch
metric learning. 1
Nishida, T., Kano, T., Goodall, J., McGrew, W. C., and
Nakamura, M. (1999). Ethogram and ethnography
of mahale chimpanzees. Anthropological Science,
107(2):141–188. 1
Pan, Y., Xu, J., Wang, M., Ye, J., Wang, F., Bai, K., and
Xu, Z. (2019). Compressing recurrent neural networks
with tensor ring for action recognition. In AAAI, vol-
ume 33, pages 4683–4690. 2
Sakib, F. and Burghardt, T. (2020). Visual recognition of
great ape behaviours in the wild. VAIB. 1, 2, 3, 4, 5
Sanakoyeu, A., Khalidov, V., McCarthy, M. S., Vedaldi, A.,
and Neverova, N. (2020). Transferring dense pose to
proximal animal classes. In CVPR, pages 5233–5242.
2, 4
Shaikh, M. B. and Chai, D. (2021). Rgb-d data-based action
recognition: A review. Sensors, 21(12):4246. 2
Sharir, G., Noy, A., and Zelnik-Manor, L. (2021). An image
is worth 16x16 words, what is a video worth? arXiv
preprint arXiv:2103.13915. 2
Simonyan, K. and Zisserman, A. (2014). Two-stream con-
volutional networks for action recognition in videos.
NeurIPS, 27. 2, 3
Triple-stream Deep Metric Learning of Great Ape Behavioural Actions
301

Tran, D., Wang, H., Torresani, L., and Feiszli, M. (2019).
Video classification with channel-separated convolu-
tional networks. In ICCV, pages 5552–5561. 2
Tuia, D., Kellenberger, B., Beery, S., Costelloe, B. R.,
Zuffi, S., Risse, B., Mathis, A., Mathis, M. W., van
Langevelde, F., Burghardt, T., et al. (2022). Perspec-
tives in machine learning for wildlife conservation.
Nature communications, 13(1):1–15. 1
Verma, V., Lamb, A., Beckham, C., Najafi, A., Courville,
A., Mitliagkas, I., and Bengio, Y. (2018). Manifold
mixup: learning better representations by interpolat-
ing hidden states. 3
Wang, L., Yuan, X., Zong, M., Ma, Y., Ji, W., Liu, M.,
and Wang, R. (2021). Multi-cue based four-stream 3d
resnets for video-based action recognition. Informa-
tion Sciences, 575:654–665. 2
Wattenberg, M., Vi
´
egas, F., and Johnson, I. (2016). How to
use t-sne effectively. Distill. 6
Zach, C., Pock, T., and Bischof, H. (2007). A duality based
approach for realtime tv-l 1 optical flow. In DAGM,
pages 214–223. Springer. 4
Zamma, K. and Matsusaka, T. (2015). Ethograms and
the diversity of behaviors, page 510–518. Cambridge
University Press. 1
Zhang, Y., Li, X., Liu, C., Shuai, B., Zhu, Y., Brattoli, B.,
Chen, H., Marsic, I., and Tighe, J. (2021a). Vidtr:
Video transformer without convolutions. In ICCV,
pages 13577–13587. 2
Zhang, Y., Wei, X.-S., Zhou, B., and Wu, J. (2021b). Bag
of tricks for long-tailed visual recognition with deep
convolutional neural networks. In AAAI, volume 35,
pages 3447–3455. 3
Zhou, B., Andonian, A., Oliva, A., and Torralba, A. (2018).
Temporal relational reasoning in videos. In ECCV,
pages 803–818. 2
Zong, M., Wang, R., Chen, X., Chen, Z., and Gong, Y.
(2021). Motion saliency based multi-stream multiplier
resnets for action recognition. Image and Vision Com-
puting, 107:104108. 2
VISAPP 2023 - 18th International Conference on Computer Vision Theory and Applications
302
