
Receptor Response and Soma Leakiness in a Simulated
Spiking Neural Controller for a Robot
David Bowes, Rod Adams, Lola Cañamero, Volker Steuber and Neil Davey
Science and Technology Research Institute, University of Hertfordshire
College Lane, Hatfield, Hertfordhsire, U.K.
Abstract. This paper investigates different models of leakiness for the soma of
a simulated spiking neural controller for a robot exhibiting negative photo-
taxis. It also investigates two models of receptor response to stimulus levels.
The results show that exponential decay of ions across the soma and of a recep-
tor response function where intensity is proportional to intensity is the best
combination for dark seeking behavior.
1 Introduction
In real neural systems it is known that leakiness in individual neurons can be caused
by a variety of physical processes which can in turn lead to a variety of temporal
profiles. Moreover the response of receptors to intensity of stimulus could be either
linear or non linear [7]. Although different mechanisms of leakiness are possible,
some are easier to implement in a robot than others. It is important to identify which
mechanisms will require the least computational effort while producing the desired
behaviour.
In this paper we investigate the effect of the specific function representing leaki-
ness and receptor response, to the ability of an artificial neural system to respond
appropriately to stimulus gradients in order to perform negative photo-taxis.
Photo-taxic robots [6], [4], [5] based on Braitenberg vehicles [3], controlled by ar-
tificial neural networks, have produced both varying results and success [6]. We have
developed a robot controller using a simplified artificial spiking neuron model, im-
plemented in an event driven simulation [8], [2]. The artificial neuron has a leaky
soma which links to an axon hillock. The axon hillock initiates spikes in one or more
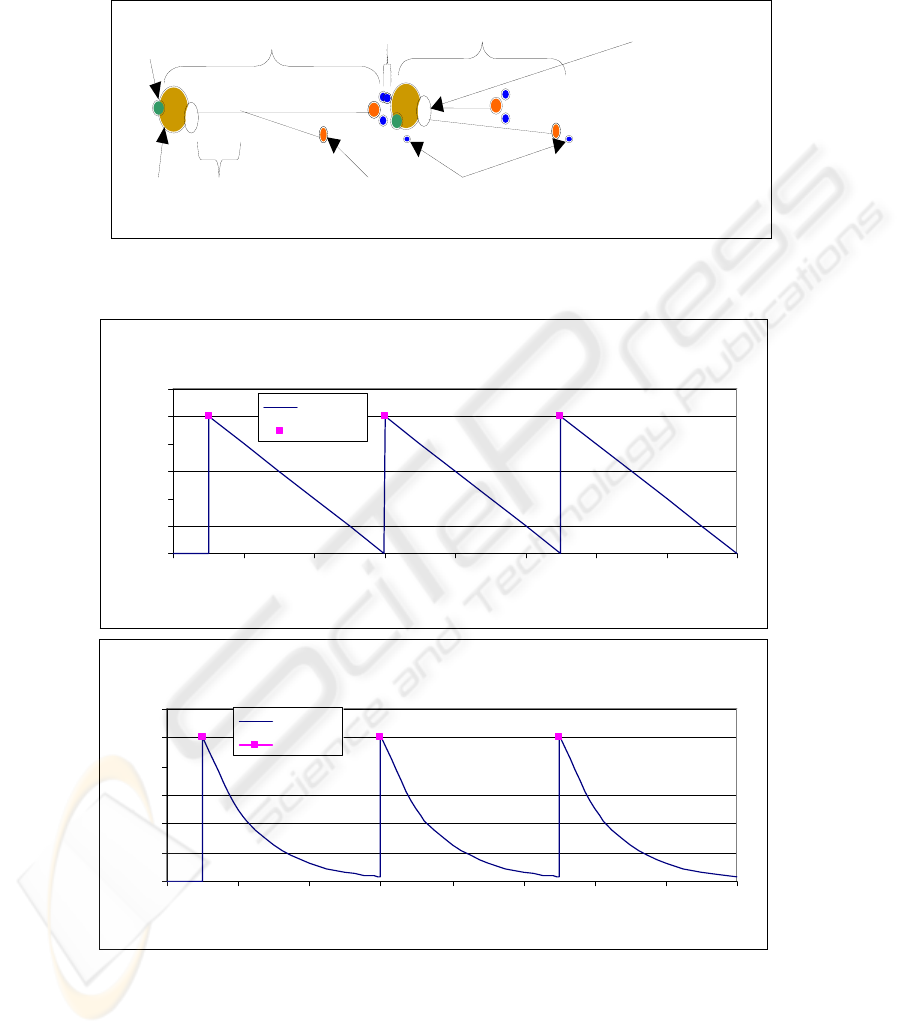
collaterals attached to the axon hillock. See Figure 1 for an illustration of the artificial
spiking neuron.
Using a modified Braitenberg fear vehicle [3], we simulated the behaviour of
negative photo-taxis. In the original Braitenberg fear vehicle the output of the left
light sensor is used to set the power output on the left motor, and a corresponding
arrangement is made on the right side of the vehicle. In our modified arrangement we
add sensory neurons, connected to interneurons, which then connect to the motor.
This gives a potentially wider range of behaviours. However this arrangement was
only satisfactory when the light intensity difference between the two light sensors was
Bowes D., Adams R., Cañamero L., Steuber V. and Davey N. (2008).
Receptor Response and Soma Leakiness in a Simulated Spiking Neural Controller for a Robot.
In Proceedings of the 4th International Workshop on Artificial Neural Networks and Intelligent Information Processing, pages 100-106
DOI: 10.5220/0001509001000106
Copyright
c
SciTePress
large. When the differences were not large, the robot was unable to correctly orientate
itself towards darkness.
Fig. 1. The model of a simplified spiking neuron showing soma, axons, chemical receptor,
synapse and neurotransmitter (note that the dendritic tree is not modeled).
Linear Decay
0
2
4
6
8
10
12
0246810121416
Time
Potential
Potential
Addition
Exponential Decay
0
2
4
6
8
10
12
0 2 4 6 8 10 12 14 16
Time
Potential
Potential
Addition
Fig. 2. Membrane potential with different decay functions with a constant rate of voltage addi-
tions generated by the arrival of spike events (designated by Addition points in the graph).
Chemi-
receptor
N
euron
1
Synapse
Soma
Axo
S
y
na
p
tic bouton
Neurotransmitter
Neuro 2
Axon hillock
101

Two aspects of the model that could account for this lack of sensitivity were thought
to be: the nature of the leakiness of the soma and the relationship between light inten-
sity and firing rate of the light receptors.
In real neurons leakiness is controlled by a variety of mechanisms including volt-
age dependent and independent ion channels, active transport though pumps, ex-
changes and axial currents [7].
Subsequently we studied two models for leakiness: linear decay in voltage, which
simple, and an exponential decay which is more plausible and could be caused by
ions passing through pores along a concentration gradient, see Figure 2.
Both of these are biologically possible, but the latter system, which does not involve
the direct use of energy, is more likely in a biological organism where parsimony of
design is often found.
The relationship between light intensity and input current to a receptor neuron is
complex and may lead to a variety of mappings between light intensity and the firing
rate of a receptor.
To investigate this, two models for firing rate of a receptor cell, under different
light intensities, were studied: inter spike time being inversely proportional to light
intensity, or, inter spike time being proportional to the difference between the maxi-
mum possible intensity and actual intensity. The first of these produces a linear rela-
tionship between light intensity and firing rate and the second produces a hyperbolic
relationship. Both of these are biologically plausible [7]. The first model would result
in firing rate being proportional to light intensity and the second method would pro-
duce a non-linear relationship as shown in Figure 3.
Firing rate v Light intensity
0
10
20
30
40
50
60
0 20406080100
Light intensity as percentage of max
Firing rate (spikes/s)
Inverse inter spike time
Linear inter spike time
Fig. 3. Firing rates of the sensory neuron using a linear and hyperbolic relation. Note the poor
discrimination of the hyperbolic function at low light intensities.).
102
2 Methodology
The experiment to test which function produced the greatest rotation was carried out
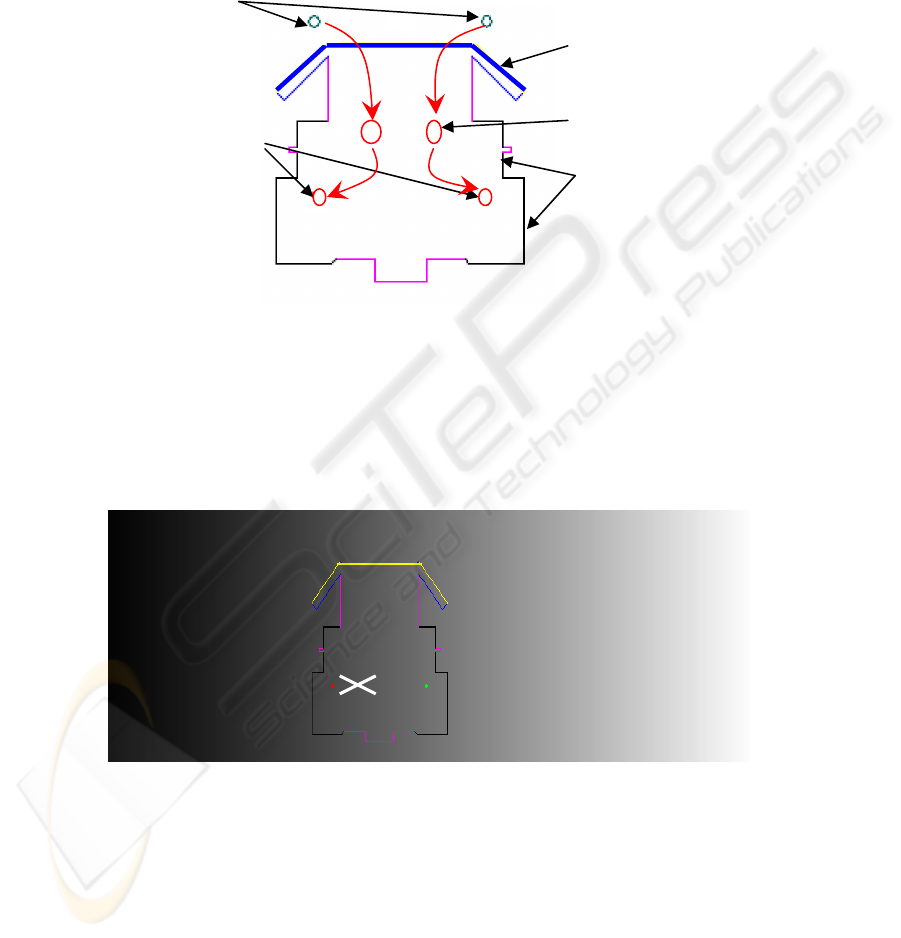
on a robot, simulated in software. A schematic of the robot is shown in Figure 4.
Fig. 4. A schematic of the robot indicating the position of the two light sensors and two in-
terneurons which attach to the motors. Note the connections are excitatory which causes the
vehicle to perform negative photo-taxis.
Simulated receptor neurons were attached to light sensors. These sensory neurons
then connected to interneurons which subsequently attached to the motors with exci-
tatory ipsilateral connections (left receptor connects to left motor). At the start of each
experiment the robot was placed perpendicular to a linear light gradient, see Figure 5.
0 Distance from black : Proportional to light intensity : Arbitrary units 10000
Fig. 5. The Robot on the linear light gradient. The robot is placed orthogonally to the gradient
and is “pinned” in position so that it can only rotate (X marks the vertical axis of rotation).
The simulation prevented forward motion of the robot, but allowed rotation so that
the robot would spin on the spot (similar to an insect being tested for pheromone
orientation in a wind tunnel). The simulation was run for 10 seconds and the total
Li
g
ht sensors
Moto
r
Wheels
Driven together
Bumper
Inter neuron
103

angle of rotation was recorded. The robot was placed at incremental distances from
the left side of the gradient, providing 9 data samples. Each permutation of decay
model, receptor firing model and position was repeated 7 times, resulting in a total of
252 experiments.
The two models for potential decay in the soma of the interneuron are:
• Linear decay: which is very simple to calculate
α
−=
+ tt
cc
)1(
• Exponential decay: biologically more plausible
β
tt
cc =
+ )1(
Where c is the synaptic concentration with constants α and β, where β < 1.
Two models for determining the time between firings of light receptor neurons
were:
• Inverse
min
1
T
i
T +∝
so
iF
∝
producing a linear relationship between light intensity and neuron firing rate.
• Linear
(
)
min
TikT
+
−
∝
so
ik
F
−
∝
1
producing a hyperbolic relationship between light intensity and firing rate.
Where k is the maximum stimulus level achievable, i is the intensity of stimulation
F is the firing rate and
T
min
is a constant which prevents the period being less than
0.01s (below this value the amount of processing increases significantly).
3 Results
The full results for all four experimental conditions are given in Figure 6. This shows
that significant rotation only occurs under the condition of exponential voltage decay
and inverse inter spike time and then only in the dark region.
A study of the motor logs for all conditions indicated that at high light levels, both
motors had been turned on to the maximum level due to very high rates of input fir-
ings. This had occurred because the soma of the interneuron had saturated and had
prevented the voltage dropping below the threshold value for spiking.
104
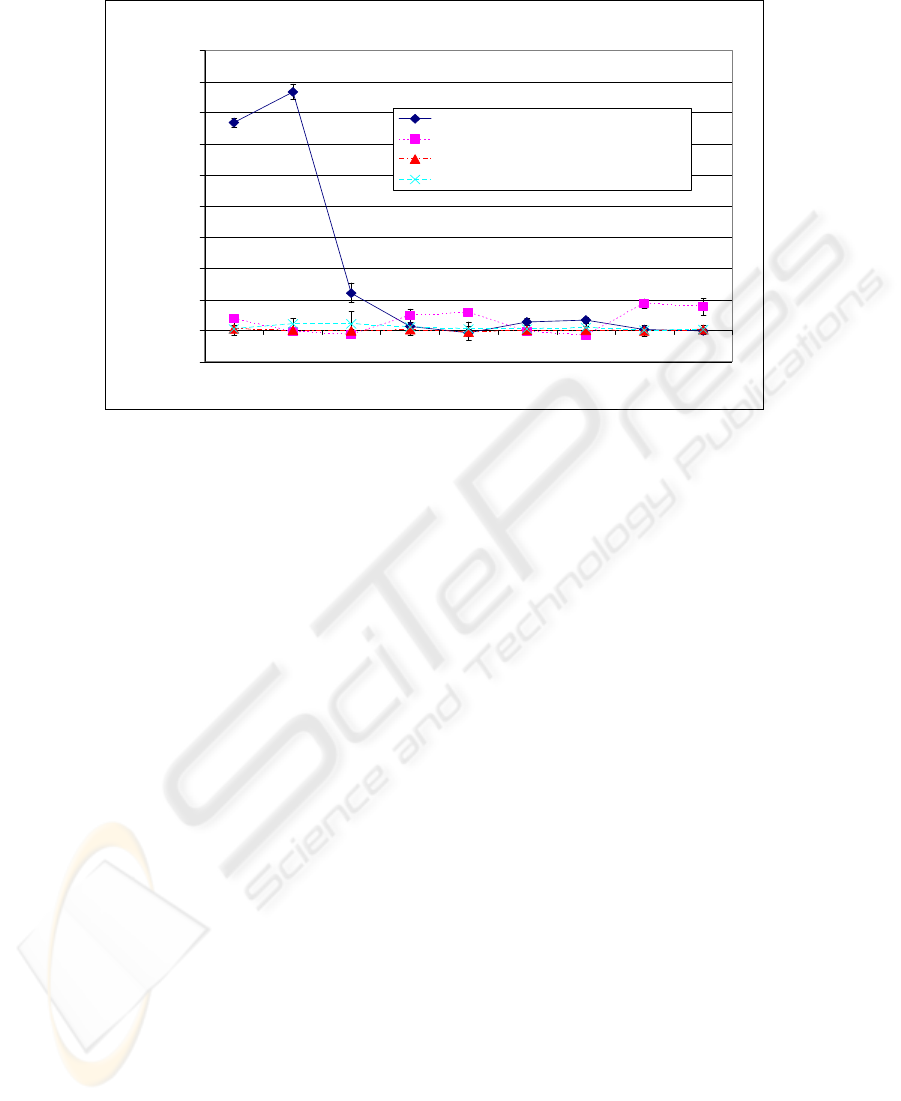
Amount of rotation for different light intensities
-0.2
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
0 1000 2000 3000 4000 5000 6000 7000 8000 9000
Distance from Black ( proportional to light intensity)
Accumulated angle of rotation (radians
)
Exp Decay, Inverse Inter Spike Time
Exp Decay, Linear Inter Spike Time
Linear Decay, Inverse Inter Spike Time
Linear Decay, Linear Inter Spike Time
Fig. 6. A graph of the amount of rotation achieved. This shows the results for the four condi-
tions: exponential voltage decay and inverse inter spike time, exponential voltage decay and
linear inter spike time, linear voltage decay and inverse inter spike time, and linear decay and
linear inter spike time. The exponential voltage decay and inverse inter spike time is the only
configuration where significant rotation occurred.
Linear decay of potential performed particularly poorly. In high light intensity the
decay of voltage in the interneuron was not sufficient to allow the voltage to fall be-
low the firing threshold – the rate of decay in this model is constant and is therefore
not related to the actual voltage. Moreover at low light intensities the voltage decay
was too rapid to allow the voltage to ever exceed the firing threshold. Exponential
decay of potential was much better: in this case the rate of decay is proportional to
voltage, producing much greater sensitivity at the interneuron.
As already noted a linear relationship between inter spike times and light intensity
at the light receptor neurons gives rise to a hyperbolic relationship between intensity
and firing rate. This gives poor discrimination at low light levels and the results dem-
onstrate the inadequacy of this encoding.
4 Discussion
In real neural systems it is known that leakiness in individual neurons can be caused
by a variety of physical processes, which can lead to a variety of temporal voltage
profiles. Moreover the response of receptors to intensity of stimulus could be either
linear or non linear [7].
Our results show that for the example of a negatively photo-taxic robot, an expo-
nential decay of membrane potential and an inverse relationship between light inten-
105

sity and inter spike time (resulting in a linear relationship between light intensity and
firing rate) produces the greatest rotation at low light levels.
At high light levels the rotation is limited due to both motors being stimulated at
the maximum rate because the interneurons receive levels of stimulus which prevents
the membrane potential from dropping below threshold.
Interestingly, the most effective neural model is also the most biologically plausi-
ble. Diffusion of ions through pores would naturally give rise to an exponential decay
whereas linearization of the decay would require additional voltage dependant mecha-
nisms.
The modified Braitenberg wiring using interneurons is biologically more realistic
than the original vehicle because it involves some level of processing by the interneu-
rons, but it is not sophisticated enough to cope with the full range of light levels. We
are currently investigating models of dynamic normalisation to achieve greater sensi-
tivity to small differences in light intensity.
References
1. Hodgkin, A. L. and Huxley, A. F: A quantitative description of ion currents and its applica-
tions to conduction and excitation in nerve membranes. J. Physiol. (Lond.), 117:500-544.
(1952).
2. Bowes,D.H; Adams,R.G; Cañamero,L; Davey,N: “A Simplified Spiking Neural Network
for a Robot” Technical report, Science and Technology Research Institute, University of
Hertfordshire (2007)
3. Braitenberg V: Vehicles: experiments in synthetic psychology. Elliot Sober (1984)
4. Floreano D: Evolutionary Robotics:A Short Tutorial (2005)
5. French, R.L.B.; Cañamero, L., "Introducing Neuromodulation to a Braitenberg Vehicle,"
Robotics and Automation, 2005. ICRA 2005. Proceedings of the 2005 IEEE International
Conference on , vol., no., pp. 4188-4193, 18-22 April (2005)
6. Bisset, D. L.; Vandenbergh, R. C: The Dynamics of Photo- Taxis: Applying the Agent
Environment Interaction System to a Simple Braitenberg Robot European Conference on
Artificial Life Vol 4, 327-336 (1997)
7. Kandel ER, Schwartz JH, Jessell TM: Principles of Neural Science, 4th ed. McGraw-Hill,
New York (2000)
8. Rochel, O; Martinez, D: An event-driven framework for the simulation of networks of
spiking neurons, European Symposium in Artificial Neural Networks, pp 295-300 (2003)
9. GM Sheperd: The Synaptic Organization of the Brain, 5thRev, Oxford UP ,New
York.(2004)
106
