
Modeling “Info-chemical” Mediated Ecological System by using Multi
Agent System
Yasuhiro Suzuki and Megumi Sakai
Department of Complex Systems Science, Graduate School of Information Science, Nagoya University,
Furocho Chikusa, Nagoya City, Japan
Keywords:
Multi Agent System, Simulation of Ecological Systems, Chemical Ecology, Evolution of Agent Communica-
tion Languages.
Abstract:
We model and simulate an ecological system, where each agent (plants, herbivores and carnivores) commu-
nicate with each other by using communication languages of chemical volatiles (info-chemical signals). This
info-chemical signals are produced by plants when they are suffered from feeding damage of herbivores and
natural enemy (carnivores) of the herbivores are attracted by the signal. In this ecological system, since carni-
vores learn the signal and trace it, plants try to endure the feeding damage until the population of herbivores
become large and enough herbivores can supply for the carnivores, otherwise carnivores are not to be attracted
by the signal and try to explore more valuable signal. However, it has reported that, some mutated plants
produce chemical signals soon even if there are few or no herbivores and attract carnivores (cry wolf plants).
We model the ecological system which cry wolf plants by using the MAS. without geographic space and with
geographic space. And we confirm that in the both types of models, in order to escape from cry wolf plants,
“honest plants” produce different types of signals so various types of signals emerge. Interestingly, in the
system with geographic space, if there is a “colony” of cry wolf plants then signal does not evolve and honest
signal and cry wolf signal can coexist.
1 CHEMICAL ECOLOGY,
ECOLOGICAL SYSTEMS WITH
INFO-CHEMICAL SIGNALS
In the science of (theoretical) Ecology, plants have
been considered as “resources” of the food chain in
the ecological system, where herbivores feed plants
and carnivores feed herbivores. Hence ecological sys-
tems have been described as interactions between her-
bivores and carnivores such as Lotoka-Volterra model
(Lotoka, 1910);
˙
X = aX − bXY, (1)
˙
Y = bXY − cY, (2)
where X is the population of herbivores and Y, carni-
vores.
However in the Chemical Ecology (Dicke and
Takken, 2008), it has turned out that plants are not
just resources but participating in the ecological sys-
tem as “a player”; they cannot run away or remove
herbivores directly but they can indirectly protects
themselves by attracting herbivores’ natural enemy
(carnivores) by producing signals; when plants are
suffered from feeding damage, they produce volatile
chemicals as “SOS signal” and attract natural ene-
mies. Plants are able to produces about twenty kinds
of signals properly and attract appropriate natural en-
emy according to the types of herbivores (Dicke and
Takken, 2008). Carnivores learn the signal and ex-
plore herbivores by tracing the signals. Hence plants
endure the feeding damage until enough herbivores
can supply for the carnivores otherwise carnivores
learn that the signal is not useful and they are not to be
attracted by the signal and explore more valuable sig-
nal (Dicke and Takken, 2008). In contrast, it has been
reported that some mutant plants produces ’cry wolf’
signals; such mutant plants produce large amount of
chemicals when they suffered small feeding damage
by a few herbivores. Such cry wolf plants produce the
same chemical as honest plant producing chemicals
(Shiojiri et al., 2010).
Shiojiri et. al. reported that “the composition
of the herbivore induced volatiles show little change
with the number of herbivores inflicting damage to the
plant, but the amounts of volatiles and the responses
of the enemies increase significantly. Signal quan-
318
Suzuki Y. and Sakai M..
Modeling “Info-chemical” Mediated Ecological System by using Multi Agent System.
DOI: 10.5220/0004332803180323
In Proceedings of the 5th International Conference on Agents and Artificial Intelligence (ICAART-2013), pages 318-323
ISBN: 978-989-8565-38-9
Copyright
c
2013 SCITEPRESS (Science and Technology Publications, Lda.)

tity may therefore provide information to the enemies
about herbivore abundance.” so such plant signals is
called ’honest’ signal (Shiojiri et al., 2010).”
In the case when most of the plants emit hon-
est signal, mutant plants (cry wolf plants) can gain
protection from herb-ivory, because visited carnivores
will remove herbivores already present. However the
number of herbivores in the cry wolf signaler are a
few for carnivores they learn such signal is not use-
ful. As (Sabelis et al., 2011) point out, when a honest
signal has been commonly used, cry wolf plants can
take advantages to use the signal and may increase its
population. However, carnivores will learn the signal
as “dis-honest” signal and carnivores may not be at-
tracted by the signal very much. So in order to keep
on attracting carnivores, honest plants haveto produce
“new-and-honest” signals. Some theoretical models
predict that co-evolution of signals will emerge in this
system (Sabelis et al., 2011; Jansen and van Baalen,
2003).
2 METHOD OF MODELING AND
SIMULATION
We model and simulate the ecological system by us-
ing two types of systems; system without / with geo-
logical space. In the system without geological space,
we regard the system as “chemical reactions,” where
each agent is chemical substance and reacts with each
other by reaction rule; such a MAS is called the Artifi-
cial Chemistry (Dittrich et al., 2006). And the system
with geological space is composed of the two dimen-
sional Cellular Automaton.
2.1 Artificial Chemistry, Abstract
Rewriting System on Multisets,
ARMS
Formally, an Artificial Chemistry, AC can be defined
by a triple (A, R), where A is the set of molecules, R
is a set of reaction rules representing the interaction
among the molecules (Dittrich et al., 2006). Various
ACs have been proposed and applied in modeling and
simulation of complex systems (Dittrich et al., 2006).
In this study, we use a model of AC, Abstract Rewrit-
ing System on Multisets, ARMS (Suzuki et al., 1996).
ARMS have been used on modeling and simulation of
non-linear chemical reactions (Suzuki et al., 2001),
Systems Biology of the cell (Suzuki et al., 2001), a
model of Origin of Life (Suzuki et al., 2001), Social
Systems and so on.
In ARMS a chemical solution is a finite multi-
set of elements denoted by symbols from a given al-
phabet, A = {a, b, . . . , }; these elements correspond to
molecules. A reaction rule is a pair of two multisets,
l, r and it rewrites a multiset, mc; if the multiset l is in-
cluded by the multiset then l is removed from the mc
and r is merged with it, so a rewriting is expressed as
mc− l + r, where “+ and -” are addition and deletion
on a multiset.
2.2 How does ARMS Work?
Example. We give R = { a,a,a → c : r
1
, b → d :
r
2
, c → e : r
3
, d → f, f : r
4
, f → g : r
5
}. and assume
that set the initial state as {a, a, b, a}. In this example,
the reaction rules are applied in parallel (the strategy
of applying rules is not only limited in parallel but
also rules can be applied rules in sequential or maxi-
mally parallel). Figure 1 illustrates a sequence of re-
{a, b, a, a} ⊆ a, b
↓ (r
1
, r
2
)
{c, d} ⊆ c, d (the left hand side of r
3
, r
4
)
↓ (r
3
, r
4
)
{e, f, f} ⊆ f (the left hand side of r
5
)
↓ (r
5
)
{e, h, h} There are no rules to apply, so it reaches
the halt state
Figure 1: Example of ARMS reaction steps
action steps, For {a, a, b, a}, r
1
and r
2
are applied and
it is transformed into {c, d} and r
3
and r
4
rewrite it to
{e, f, f }; no rules can rewrite it so rewriting halts.
3 MODEL WITHOUT
GEOGRAPHIC SPACE
It has been reported that every honest plant produces
almost same quantity of HIPV and threshold of the
population of herbivores for start producing HIPV
almost the same (Dicke and Takken, 2008; Shiojiri
et al., 2010). Hence we can estimate the degree by
feeding damage by herbivores based on the quantity
of the HIPVs. So we define the biomass of plants,
P as P− ΣH
i
, where H
i
is the total quantity of every
HIPVs; if the quantity of HIPVs are small, there are
few herbivore and, large quantity of HIPVs indicates
there are many herbivores.
The model is composed of honest signal, h
i
, dis-
honest signal, c
i
and a carnivore which is attracted
by h
i
or w
i
; where the suffix “i” indicates the type of
chemical profile of HIPV. Plants produce h
i
and the
carnivore is attracted and remove herbivores. When
Modeling"Info-chemical"MediatedEcologicalSystembyusingMultiAgentSystem
319
the population of honest signal increased, dis-honest
signal of it emerges and if a carnivore is attracted
by the dis-honest signal, it learns the signal is dis-
honest and does not to be attracted by the honest sig-
nal. Hence, if the dis-honest signals increased, hon-
est plant produces different chemical profile of HIPV.
Honest plants can not know, how manydis-honest sig-
nals are produced, however if they can not produce
different type of HIPV and attract carnivores, they
may go extinct. Hence, if the honest plants survive
under the large quantity of dis-honest signals, it indi-
cates that honest plants can produce different type of
HIPV. On the other hand, when the quantity of dis-
honest signals is small, if a honest plant produces dif-
ferent type of HIPV, carnivores may not to be attracted
to the new HIPV, because they do not have to learn it
is dis-honest signal (in fact, it is not dis-honest signal
because of the population of dis-honest signal is low).
Hence when the the quantity of dis-honest signal
is large, if the honest plant using the same chemi-
cal profile as dis-honest signal is mutated and can
produce new honest signal and attract carnivores in-
creases its population, therefore it can be regarded as
if the population of dis-honest signal increases, there
emerges new honest signal. In this model, the proba-
bility of emergence of new HIPV is defined as
Σh
i
Σ(h
i
+ w
i
)
> τ,
where h
i
indicates the honest signal of type i and w
i
,
the dis-honest signal of type i and τ is the threshold
value of emerging the new type of HIPV.
The model is composed of following reaction
rules;
h
i
k
1
→ h
i
, h
i
(r
1
),
h
i
k
3
→ c
i
, h(r
2
),
c
i
, h
i
k
2
→ c
i
(r
3
),
h
i
k
5
→ w
i
, h
i
(r
4
),
c
i
, w
i
k
6
→ w
i
(r
5
),
w
i
k
7
→ ⊥(r
6
),
c
i
k
4
→ ⊥(r
7
),
where r
1
expresses the population growth of herbi-
vores; as the population of herbivores is increased, the
quantity of HIPV is also increased. r
2
expresses the
HIPV attracts carnivores and r
3
expresses attracted
carnivores remove herbivores. r
4
expresses mutation
of plants from honest to cry wolf plant and r
5
ex-
presses the learning of carnivores; they learnt and
avoid to be attracted by the HIPV so the population
of carnivores go to the HIPV is decreased. r
6
and r
7
are natural death of dis-honest plants and carnivores.
3.1 Result of Simulation
We set the reaction constants, k
1
to k
7
as 0.9, 0.5, 0.5,
0.4, 0.5, 0.5, 0.5, respectively, τ = 0.6 and the quantity
of honest signal is 30, the quantity of dis-honest sig-
nals and the number of carnivores are zero, in the ini-
tial state; this parameter setting indicates the growth
of herbivore is high and the mutation rate from honest
plant to wolf is small. Since the quantity of dis-honest
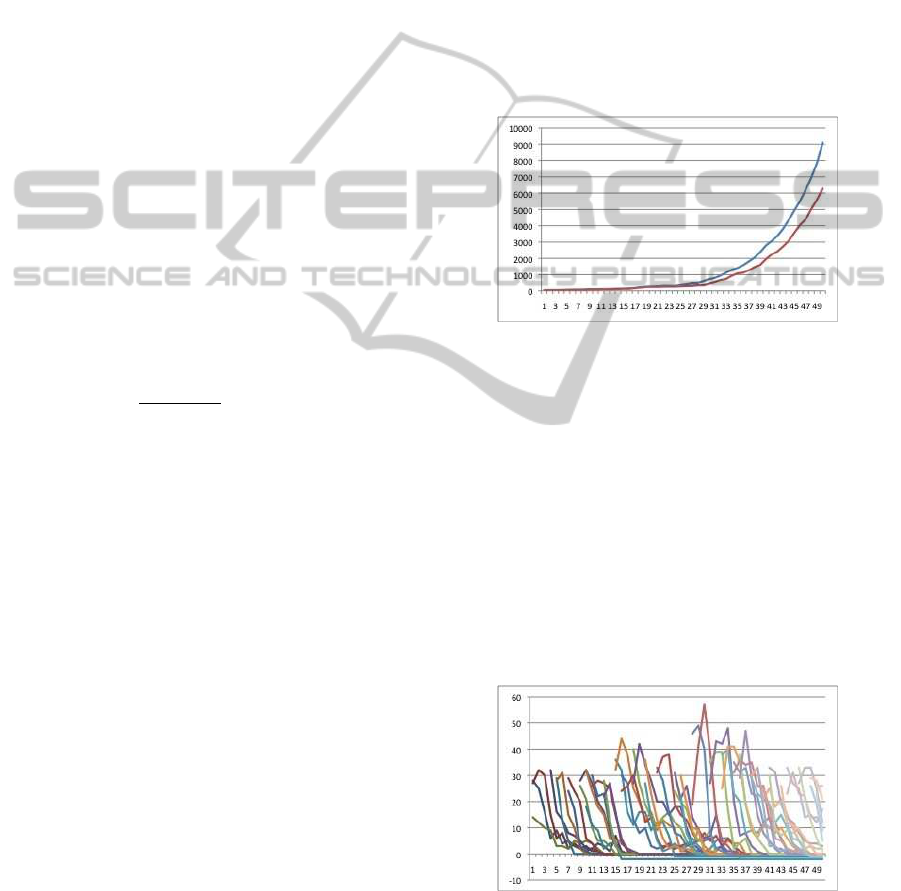
signal is not too large, HIPV does not evolve (Fig-
ure 2). Next, we increase the quantity of dis-honest
Figure 2: When the quantity of dis-honest signals are not
too large, upper line indicates the quantity of honest sig-
nal and below, dis-honest signal; the vertical axis illustrates
the quantity of HIPV (both honest and dis-honest signals)
and the horizontal axis illustrates the steps. The biomass of
plant is 10,000 so the plant biomass reaches to near zero
signals and retarded carnivores attractiveness to the
HIPV, in order to realize it, we change the reaction
constants for the rule of w
i
, c
i
→ w
i
(r
5
). We set k
1
to k
7
as 0.9, 0.5, 0.5, 0.7, 0.5, 0.5, 0.5, respectively,
τ = 0.6 and the quantity of honest signal is 30 and
quantity of dis-honest signals and the number of car-
nivores are zero, in the initial state. In this case, HIPV
evolved and various types of HIPV emerge (Figure 3).
Figure 3: When the quantity of dis-honest signals are large,
each lines indicates the quantity of honest and dis-honest
signals; the horizontal axis illustrates the quantity of HIPV
(both honest and dis-honest signals) and the horizontal axis
illustrates the steps.
ICAART2013-InternationalConferenceonAgentsandArtificialIntelligence
320

4 MODEL WITH GEOGRAPHIC
SPACE
We model the system by using two dimensional Cel-
lular Automaton (2DCA); the geographic space is
square lattice and the eight cells surrounding a cen-
tral cell (Moore neighbour); in the initial state, plants
are distributed on the lattice; every plant grows in the
same growth rate and when the its size (we call the
size of a plant as “biomass” in the below) reaches the
given value, stop growing and a new plant sprout in
the neighbouring empty space.
In several randomly selected plants, herbivores
come and start feeding; they increase the population
by feeding and if the number of herbivoresexceeds its
biomass, the plant goes to die out and all herbivores
move and distribute randomly to the nearest plants;
plants start producing chemical signal when the popu-
lation of herbivores exceeds the given threshold (hon-
est plant). Each carnivores search the chemical signal
by walking on the lattice, carnivores expense “phys-
ical power” by walking and if a carnivore finds the
plant which has been suffered from feeding damage
by herbivores (we call such a plant as “patch” in
the below) , the carnivore removes all herbivores and
gains its physical power according to the number of
herbivores in the plant, while if a carnivore spends all
physical power before finding the plant having feed-
ing damage, it goes to die.
4.1 Cry Wolf Plant and Evolution of
Signal
When a new plant sprouts, randomly selected plant
become “cry wolf plant” and it generates chemical
signal when it is suffered from small amount of feed-
ing damage; in the initial state, there are no cry wolf
plants and all plants and carnivores uses the same sig-
nal; however in the lapse of steps, the number of cry
wolf plants increase and carnivores attract to such cry
wolf plants. Each carnivore judges the quality of sig-
nal by “(the number of herbivores in the plant) / (the
distance from the former patch) = Evaluation Value
(EV)”, if the EV is lower than given threshold, the
carnivore judge and learn the plant as cry wolf and
attractiveness of the signal becomes low. Hence as
the population of cry wolf plants increases, the attrac-
tiveness of carnivores becomes low, so honest plants
have to produce different HIPV. In ecological sys-
tems, different HIPVs are continuously generated by
mutated plants (Shiojiri et al., 2010), if there are no
cry wolf plants, carnivores do not learn other HIPV
and plants producing different type HIPV can not at-
tract carnivores and die out, however if the population
of cry wolf plants increases, carnivores explore differ-
ent HIPV randomly so such mutated plants can have
the possibility to attract carnivores.
Learning of a Carnivore
In order to implement learning activity of a carnivore,
we use the simple stochastic model, the polya’s urn;
this model is composed of a urn (pot) and balls, balls
are painted with some sorts of colors. In this model,
history of picking up a ball is remembered through
following way; let us assume that there are three types
(colors, red, black, white) of balls in a pot and the
number of each colored balls are same (for simplicity,
we assume that there are same number of each colored
balls in the urn, here); when a “black” ball is picked
up from the pot, n, (n > 0) black balls are returned to
the urn, hence the probability of selecting a black ball
is increased, compared with other colors. We express
the type of HIPV as the color of ball and the popu-
lation of each HIPV as the number of balls. When
a carnivore judges a HIPV as “honest signal” n balls
of HIPV are added to its memory (urn) and if judges
as “cry wolf”, m balls of HIPV are removed from the
memory and the m balls of “random walk” are added
instead of them; when the carnivore select the ball of
random walk, it explore nearest HIPV randomly.
5 RESULT OF SIMULATION
In the model with geological space, when the popu-
lation of cry wolf plants increase, carnivores explore
different type of HIPV and HIPV evolves (Figure.4).
However, as the types of HIPV becomes larger, the
Figure 4: The time evolution of population of HIPV; each
lines illustrates different HIPV.
number of plants producing each HIPV become rela-
tively low; for example when there are 100 plants and
produce two types of HIPV, if 50 plants produce the
same HIPV, the probability of finding each of HIPV
is 50/100, however if each of plant produces differ-
ent HIPV, the probability of finding each of HIPV be-
comes 1/100; so as the types of HIPV increase, the
Modeling"Info-chemical"MediatedEcologicalSystembyusingMultiAgentSystem
321

most of carnivores expense all physical power before
finding out the target HIPV producing plant and they
go extinct. We confirmed this phenomenon in this
model; this phenomenon is called the “Tower of Ba-
bel” and several theoretical contributions have been
addressed it (Jansen and van Baalen, 2003). Hence,
cry wolf plants may induce “Tower of Babel” effect,
but we discover that when they form a “patch” in
the geographic space, HIPV does not evolve and the
Tower of Babel does not emerge (Figure5).
Food Court in an Ecological System
When cry wolf plants are sparsely distributed, carni-
vores may move long distance and meet a cry wolf
plant, it expenses physical power largely but can not
regain the power very much, because there are few
herbivores and it does not enough for long traveled
carnivore; However, if cry wolf plants form a patch,
carnivores do not have to travel long distance and just
hop around cry wolf plants and obtain herbivores;
even if the population of herbivores in each plant is
small, if it is a large patch, it will be able to offer
enough herbivores to carnivores as a whole of the
patch.
And honest plants do not produce HIPV soon, so
carnivores have to wait until population of herbivores
increases, but cry wolf plants produce HIPV soon
and they can offer small amount of herbivores; so to
speak, honest plants serve “full course meal” and cry
wolf plants serve “light meal”; a patch of cry wolf
plants is regarded as a “food court of light meals”
hence carnivores do not judge the HIPV of cry wolf
plants forming a patch as “honest signal” and HIPV
does not evolve (Figure 5). In simulations, we use
the same number of cry wolf plants and compare the
case when they are distributed randomly with placed
gathered; and confirm that in the former case, HIPV
evolves and in the latter case, HIPV does not evolve.
Figure 5: Time evolution of HIPV; each lines illustrates the
different type of HIPV; there are several HIPVs in low con-
centration, they emerges by mutation of plants and changes
of concentration are due to the growth and feeding damage
by herbivores; even if such plant produces HIPV, carnivores
do not learn other HIPV and they can not attract carnivores
and die out
6 DISCUSSION
In order to consider the effect of geographic structure
by using a model without using geographic structure,
we consider a simple example. Let us consider a sim-
ple ARMS: the initial state is {a} and it has a rule
R = {a → a, a}; if we consider this reaction by using
the differential equation,
a
dt
= a,
this is the Malthus equation and the number of a in-
creases exponentially. However, in this ARMS the
rule is applied sequentially (one rule is applied in the
each step time), the time evolution of computation
which starts from {a} is;
{a} → { a, a} → {a, a, a} → . . . ,
hence the increment of the number of a is described
as
M(a
i
) = M(a
i
1
) + 1,
where M(a
i
) denotes the multiplicity of a at step time
of i; and the number of a increases lineally.
If the rule is applied in maximally parallel, the
rule is applied as much as possible in the each step,
the time evolution is {a} → { a, a} → {a, a, a, a} →→
{a, a, a, a, a, a, a, a, a, a, a, a, a, a, a, a} → . . . , so the
increment of the number of a is described as
M(a
i
) = 2 × M(a
i−1
),
and the number of a increases exponential (non-
linearly), 2
i
, (i = 0, 1, 2, ..., ) and time evolution fits
with the Malthus equation.
6.1 Linear, Non-linear and
“Meso”-linear
In the ARMS with the maximally parallel rule appli-
cation, reaction constants are defined for each reac-
tion rule. For example, if we set the reaction con-
stant for the rule a → a, a as 0.5, the expectation value
of applying the rule is 0.5 × the size (cardinality)
of the multiset; so the time evolution is, for exam-
ple {a} → {a, a} → {a, a, a, a} → {a, a, a, a, a, a} →
{a, a, a, a, a, a, a, a, a, }. . ., where the rule is applied in
parallel to bold characters. Hence the time evolution
is neither linear nor non-linear. We namesuch dynam-
ics as “meso”-linear, which means that the dynamics
is in between linear and non-linear. In this example,
when the reaction constant near to 1.0, its behavior
resembles with the Malthus equation and near to 0.0,
resembles linear system.
ICAART2013-InternationalConferenceonAgentsandArtificialIntelligence
322
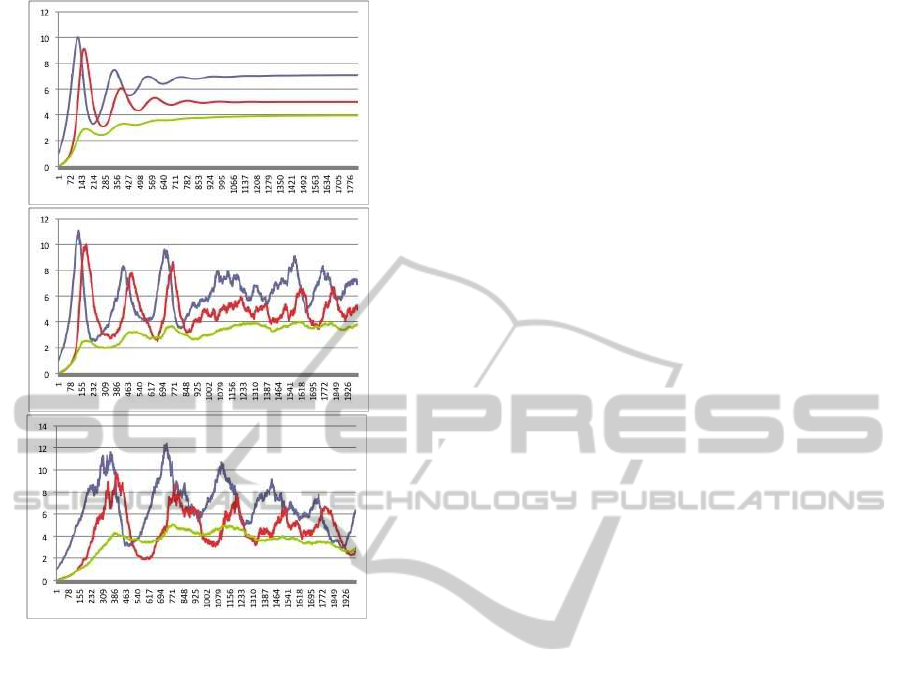
Figure 6: Simulation of the rate equation of the model of
the Chemical Ecological system in the Section 3; the top is
the same as Differential Eauation and from top to bottom,
non-linearity becomes strong.
6.1.1 System Size and Meso-linearity
If the size of multiset and the reaction constant are
small, the number of applicable rules is to be also
few, on the other hand, if the size of multiset is large,
even if the reaction constant is small, the number of
applicable rules are not so few. In the ARMS with
the rule of a → a, a, the size of multiset in the initial
state is more than 10, each of time evolution is not
so different, however, when the size of multiset is 1
({a}), its time evolution is different from others; we
investigated the time evolution by changing the reac-
tion constants as 1.0, 0.9, 0.8, ..., 0.1 and the size of
multiset in the initial state as 10,000, 1,000, 100, 10,
1 and examine the meso-linearly compared with the
reaction constant is 1.0 (maximally parallel);
We investigate the time evolution of the popula-
tion of a in 10 steps from the initial state, the rule is
applied in maximally parallel with reaction constants
as 1.0, 0.9, 0.8, ... 0.1.
In each step, the difference between the popula-
tion a with the reaction constant is 1.0 and others and
the difference is divided by the size of multiset in the
initial state for normalizing the value; so we confirm
that when the size of multiset is 1, its time evolution is
different from others. We transform the reaction rule
of the model of chemical ecology (r
1
to r
7
in the Sec-
tion 3) into the rate equation of chemical reactions and
we confirmed that by inducing meso-linearlity, behav-
ior of time evolutions becomes different (Figure6).
Considering relation between meso-linearity and the
effect of geological space is our future work.
ACKNOWLEDGEMENTS
This work was supported by the JSPS Core-to-
Core Program (No.20004), the MEXT Grant-in-Aid
for Scientific Research on Innovative Areas (No.
24104002) and Grant-in-Aid for Scientific Research
(No. 23300317).
REFERENCES
Dicke, M. and Takken, W. (2008). Chemical Ecology.
Springer Verlag.
Dittrich, P., Ziegler, J., and Banzhaf, W. (2006). Artificial
chemistries - a review. Artificial Life, 3(7):225–275.
Jansen, V. and van Baalen, M. (2003). Common language
or tower of babel? on the evolutionary dynamics of
signals and their meanings. Proceedings of Biological
Science, 7(270):69–76.
Lotoka, A. J. (1910). Contribution to the theory of periodic
reaction. Journal of Physical Chemistry, 14(3):271–
274.
Sabelis, M. W., Janassen, A., and Takabayashi, J. (2011).
Can plants evolve stable alliances with the enemies’
enemies? Journal of Plant Interactions, 2(2-3):71–
75.
Shiojiri, K., Ozawa, R., Kugimiya, S., Uefune, M.,
Wijk, M., Sabelis, M., and Takabayashi, J. (2010).
Herbivore-specific, density-dependent induction of
plant volatiles: honest or cry wolf. PLoS One,
17(5(8)):e12161.
Suzuki, Y., Fujiwara, Y., Takabayashi, J., and Tanaka, H.
(2001). Artificial life application of p systems. In
LNCS, volume 2235, pages 299–346. Springer Verlag.
Suzuki, Y., Tsumoto, S., and Tanaka, H. (1996). Analysis
of cycles in symbolic chemical system based on ab-
stract rewriting system on multisets. In Artificial Life
V, pages 521–528. MIT Press.
Modeling"Info-chemical"MediatedEcologicalSystembyusingMultiAgentSystem
323
