
AN IDIOTYPIC NETWORK APPROACH TO TASK
ALLOCATION IN THE MULTI-ROBOT DOMAIN
Use of an Artificial Immune System to Moderate the Greedy Solution
Amanda Whitbrook
1
, Gabriel Gainham
2
and Wen-Hua Chen
2
1
BAE Systems, Systems Engineering Innovation Centre (SEIC), Loughborough University
Loughborough, Leicestershire, LE11 3TU, U.K.
2
Department of Aeronautical and Automative Engineering, Loughborough University
Loughborough, Leicestershire, LE11 3TU, U.K.
Keywords: Multi-Robot Task Allocation (MRTA), Artificial Immune System (AIS), Idiotypic Network, Autonomous
Mine Clearance.
Abstract: This paper presents and explains a set of equations for governing simultaneous task allocation in multi-robot
systems and describes how they are used to construct a novel algorithm - the Idiotypic Task Allocation
Algorithm (ITAA); the equations are based on Farmer's model of an idiotypic immune network but are
adapted to include 2-dimensional stimulation and suppression and the use of affinity rather than
concentration levels to select antibodies. This novel approach is taken to render the model suitable for
simultaneous task allocation where robots must act individually; other idiotypic algorithms have only been
applicable to problems where many robots are required to perform one task at a time using swarming
behaviours. The paper describes the analogy between idiotypic network theory and the problem of task
allocation and shows how the former can be used to increase the fitness of solutions to the latter, also
discussing the types of Multi-Robot Task Allocation (MRTA) problem that might benefit from this
approach. The results of applying ITTA to a number of simulated mine-clearance problems (with increasing
numbers of robots and mines) are presented, and clear advantage over the greedy solution in both simple
and more complex scenarios is demonstrated.
1 INTRODUCTION
There are many different types of Multi-Robot Task
Allocation (MRTA) problem including varying
combinations of single-task (ST) robots, multi-task
(MT) robots, single-robot (SR) tasks, multi-robot
(MR) tasks, instantaneous assignments (IA, with no
planning for future allocations), time-extended
assignments (TA, which allows for future allocation
planning) and online assignment variations of IA
(OA, where tasks are introduced one at a time). The
interested reader is directed to Gerkey and Mataric
(2004), which presents a comprehensive,
architecture-independent taxonomy. In addition,
robots may be heterogeneous in their capabilities
and performance, and tasks may differ in
complexity, difficulty and solution requirements.
Whilst all types of MRTA problem may be solved
by implementing a greedy algorithm, characterised
by repeatedly taking the 'best' valid option (based on
some measure of fitness) at a local level,
optimization is not guaranteed. In addition, the
Linear Programming (LP) approach, which
guarantees optimality, cannot be applied to some of
the more complex MRTA problem types including
ST-SR-IA-OA, ST-SR-TA, ST-MR-IA, ST-MR-TA,
MT-SR-IA, MT-SR-TA, MT-MR-IA and MT-MR-
TA combinations, some of which are strongly NP-
hard (Gerkey and Mataric (2004)). There is thus a
need for heuristic approaches that are capable of
providing fitter solutions than those offered by
greedy algorithms, and much research effort has
been directed towards the development of heuristic
MRTA techniques. For example, auction-like
allocation mechanisms are described in Guerreor
and Oliver (2011), Nanjanath and Gini (2010) and
Gerkey and Mataric (2002). There have also been a
number of works published on market-based
techniques (for example Dias et al. (2005)),
coalition-formation methods (Shehory and Kraus
5
Whitbrook A., Gainham G. and Chen W..
AN IDIOTYPIC NETWORK APPROACH TO TASK ALLOCATION IN THE MULTI-ROBOT DOMAIN - Use of an Artificial Immune System to Moderate
the Greedy Solution.
DOI: 10.5220/0003709000050014
In Proceedings of the 4th International Conference on Agents and Artificial Intelligence (ICAART-2012), pages 5-14
ISBN: 978-989-8425-96-6
Copyright
c
2012 SCITEPRESS (Science and Technology Publications, Lda.)

(1998)), self-organisation (Fukuda et al. (1988)) and
emergent systems (Liu et al. (2007) and Atay and
Bayazit (2007)).
This paper presents a set of equations based on
those developed by Farmer et al. (1986) that
represent an idiotypic immune system approach (see
Jerne (1974)) to solving the problem of task
allocation in the multi-robot domain. The equations
have been adapted to include 2-dimensional
stimulation and suppression and the use of affinity
rather than concentration levels to select antibodies.
This is a novel approach that allows each robot to
solve a separate task independently so that all tasks
can be completed simultaneously.
The paper describes the analogy between task
allocation and the idiotypic network theory of the
immune system and shows how the equations can be
applied to general ST-SR-IA problems with N robots
looking for one task to complete and L tasks
requiring one robot. It sets out how this approach
differs from previous idiotypic implementations of
MRTA and explains its advantages over them; in
particular, other idiotypic algorithms have only been
applicable to problems where many robots are
required to perform one task at a time using
swarming behaviours.
A set of experimental results on simulated
problems of this type is presented, initially where L
= N, N varies between 3 and 15, and robots are
required to organise mine diffusion tasks in a way
that minimizes travel costs. Some preliminary results
for the case where N ≠ L are also briefly discussed.
The results provide empirical evidence that the
Idiotypic Task Allocation Algorithm (ITTA)
described here is capable of outperforming the
greedy approach such that mean fitness is
significantly improved for these problem types.
2 BACKGROUND, PRIOR WORK
AND MOTIVATION
2.1 Background
The purpose of the immune system is to identify and
neutralize the molecules or cells that are dangerous
to the body (antigens) without damaging healthy
cells (Barra and Agliari (2007)). This is achieved
through the interaction of many different types of
immune cell, which each have specific roles. The
main constituents of the adaptive immune system are
B-lymphocytes (B-cells) and T-lymphocytes (T-
cells), which have particular protein molecules on
their surfaces called receptors. The receptors of B-
cells can bind to antigens that 'match' them, allowing
the B-cells to neutralize them.
The clonal selection theory of the immune
system (Burnet (1958)) states that lymphocytes
operate independently, and that once a match is
established, B-cells proliferate (increase in
concentration) by cloning and releasing free
receptors known as antibodies. Binding takes place
between a region of the antibody known as the
paratope and a region of the antigen known as the
epitope. In contrast, Jerne's idiotypic network theory
of the immune system (Jerne (1974)) postulates that
lymphocytes interact with each other so that the
immune system functions as a global network of
cells stimulated and suppressed by internal
recognition and matching between themselves. This
is because antibodies also serve as internal images of
certain antigens and are thus themselves being
detected and acted upon (Barra and Agliari (2007)),
which keeps the concentrations of antibodies at
appropriate levels. Antibody paratopes are thus not
only matched to antigen epitopes but also to epitope
regions on other antibodies, known as idiotopes.
Figure 1 below shows the structure of an antibody
and illustrates how antibody concentrations are
suppressed by other antibodies that recognise their
idiotope, and how concentrations are stimulated to
increase when they recognise another antibody's
idiotope.
Figure 1: Antibody paratope and idiotope regions.
2.2 Prior Work
The dynamics of antibody and antigen
concentrations are modelled computationally as
differential equations in Farmer et al. (1986). This
model is widely used for constructing Artificial
Immune System (AIS) implementations of idiotypic
networks, especially in navigational robotics, where
the method has demonstrated flexible behaviour-
mediation properties. However, in this field artificial
ICAART 2012 - International Conference on Agents and Artificial Intelligence
6

idiotypic networks have largely been confined to
single robot navigation problems, for example,
Watanabe et al. (1998), Vargas et al. (2003), Luh
and Liu (2004), and Whitbrook et al. (2007), where
the individual behaviours of single robots are
modelled as antibodies and environmental
information is modelled as antigens.
On the other hand, the application of idiotypic
principles to task allocation in the multi-robot
domain is somewhat more scarce, especially
utilization of the Farmer-based model, despite the
fact that its decentralized yet cooperative and
coordinated approach to problem solving lends itself
very elegantly to such systems. Sathyanath and
Sahin (2002) implement idiotypic mine detection but
use a simplistic analogy rather than the Farmer
model, i.e., idiotopes are not modelled and play no
role in determining the stimulation and suppression
levels of robots. Mitsumoto et al. (1995) implement
swarm behaviour by using a clonal selection-based
method rather than an idiotypic network; self-non-
self discrimination is modelled and tactics between
the robots are secreted and proliferated until
swarming behaviours emerge. Dioubate et al. (2008)
use a hybrid Farmer-based idiotypic network
coupled with clonal selection and genetic evolution
of lymphocytes to generate co-ordinated formation
of robots behind obstacles. Lee and Sim (1997) use
the Farmer model to develop idiotypic cooperative
strategies leading to swarm behaviours; robots
communicate their behaviours to each other on a
local level and the behaviour (antibody) that shows
the greatest stimulation is adopted by the whole
group. Jun, Lee and Sim (1999) and Sun, Lee and
Sim (2001) use an extended version of this model
that includes additional T-cell control of
concentrations to improve the adaptation capability.
Razali et al. (2009, 2010) use the same model as
Jun, Lee and Sim (1999), but also include memory
enhancement to achieve shepherding behaviour for
robot dogs managing robot sheep. Li et al. (2007)
solve the same problem, also using Farmer's
idiotypic model, but do not include T-cell control or
enhanced memory.
2.3 Motivation
In all of the examples cited above either the Farmer
model is not implemented or the goal is to adopt
majority behaviour patterns rather than assign
individual behaviours to individual tasks. The
general Lee and Sim approach is thus suited to
problems where a number of tasks that require many
robots to solve them are completed in sequence (ST-
MR-TE), but it is not applicable to the broader
spectrum of problems including those that require
instantaneous assignment (IA) of robots to different
tasks. In essence, the Lee and Sim analogy is the
same as for single robot navigation, i.e., behaviours
are modelled as antibodies, and only one behaviour
is adopted at a given time. If robots in the group are
required to adopt different behaviours at a given
time (as in IA problems), then a different model is
clearly needed. Furthermore, there is a real
requirement for IA-type assignment of
heterogeneous tasks to heterogeneous robots within
the military domain. In particular, it is envisaged
that within the next twenty-five years autonomous
military capabilities will undergo a major shift
toward joint, multi-mission, collaborative operations
between manned and unmanned vehicles (US
Department of Defence (DOD) (2009)). For
example, fleets of unmanned aerial vehicles (UAVs)
and unmanned ground vehicles (UGVs) will be
required to work together to accomplish
reconnaissance, surveillance, mine detection and
target-designation missions. Within such operations,
successful task allocation and coordination of the
many heterogeneous assets will be critical to mission
success, but will also impose a great burden on
central command and control as the number of assets
increases. For this reason, an autonomous,
decentralized, self-regulating coordination system in
which the assets are able to allocate tasks
independently of human control would be of great
value to the military. In addition, if progressed
through to use in theatre, a successful framework for
decentralized coordination and control of
heterogeneous, multi-agent, military systems would
represent a significant step forward for autonomy.
Indeed, the current US DOD Unmanned Air
Systems Roadmap (2005) cites “distributed control”
as the main criteria for achieving an autonomy level
of 8 in the DOD scale (range = 1 to 10) compared
with the remotely-operated systems that are typically
in place at present; these are measured as between
levels 1 and 3 on the same scale. This paper sets out,
describes and tests the Idiotypic Task Allocation
Algorithm (ITAA), which provides a potential
solution to the problem of autonomous,
decentralized, distributed task allocation for IA-type
assignment of heterogeneous tasks to heterogeneous
robots.
3 PROBLEM SPACE
A mine-clearance scenario has been selected as the
AN IDIOTYPIC NETWORK APPROACH TO TASK ALLOCATION IN THE MULTI-ROBOT DOMAIN - Use of an
Artificial Immune System to Moderate the Greedy Solution
7

test-bed for the ITAA as it has many properties that
make it ideal. In particular, there is sufficient
flexibility within the problem space to allow more
simple variations to be implemented in the early
stages of research, and to build and test more
complex instances as the work progresses, for
example beginning with ST-SR-IA experiments,
equal numbers of identical tasks (mines to diffuse)
and identical robots, incrementally building up to the
inclusion of online assignments (OA), time-extended
assignments (TE), unequal numbers of tasks and
robots, heterogeneous tasks and robots, multi-task
robots (MT), multi-robot tasks (MR), and real-time,
real-world implementations that require additional
features such as reactive obstacle avoidance
modules.
In this paper, research begins with the problem
of assigning a known number L of identical, un-
diffused mines to a known number N of
homogenous robots in simulation. Initially, it is
assumed that:
1. the robots have equal capabilities and travel at
the same, fixed speed;
2. the mines are equally accessible to all the
robots, i.e., there are no obstacles to negotiate;
3. the level of difficulty in diffusing a mine is
equal for all mines and constant throughout the
operation;
4. the number of mines L does not change at any
time during the operation;
5. the number of robots N does not change at any
time during the operation;
6. the number of robots available is always equal
to the number of mines needing diffusing, i.e., N
≡ L.
7. once assignment has taken place and the mines
are diffused, all work is done.
Note that assumptions 1 to 3 allow the distance
between robots and mines to be used as a measure of
affinity between them. If this were not the case, then
a more complex measure would be needed, i.e. one
that also considers the ability of each robot to
complete each individual task and the additional
time that would be needed to negotiate (possibly
moving) obstacles. This paper is chiefly concerned
with validating the theory set out in Section 4 so use
of the most simplistic case in the first instance
allows the essential theory of the ITTA model to be
tested independently of any real-world noise. The
results of a preliminary investigation into cases
where N ≠ L is also briefly discussed here but more
complex experiments will be conducted in the future
in order to establish whether the method stands up to
the problems associated with real-world
implementation.
4 THE IDIOTYPIC TASK
ALLOCATION ALGORITHM
(ITAA)
In the model presented here, antibodies are
analogous to possible robot-mine pairs, and the
affinity U of each antibody to the current antigen
(physical positioning of all robots and mines) is the
distance d between the robot and mine in the
antibody pair. This is the antibody pre-affinity,
before stimulation and suppression from other
antibodies are taken into consideration. The post-
affinity after stimulation and suppression is denoted
as T. Writing this more formally, U
ij
is the pre-
affinity, T
ij
is the post-affinity and d
ij
is the distance
between robot i, i = 1, …, N and mine j, j = 1, …, L.
The pre-affinity is thus given by
=
.
(1)
After pre-affinities have been calculated, the
initial allocation of robots to mines is achieved by
executing a simple and intuitive greedy algorithm
where the antibody with the smallest affinity is
repeatedly selected as an allocation and then all pairs
that contain that robot and mine are eliminated from
future allocations until exactly one robot is allocated
to exactly one mine. This greedy algorithm is also
known as the Sequential Best-Pair Algorithm
(SBPA, see Oliver and Guerrero (2011)). Let
represent the index of the robot allocated to mine j
based on pre-affinities. Under the ITTA model, this
is the antigenic robot to mine j, one of a set of L
antigenic robots (as exactly one robot is antigenic to
each of the L mines). Similarly, let represent the
index of the mine allocated to robot i based on pre-
affinities. This is the antigenic mine to robot i, one
of a set of N antigenic mines (as exactly one mine is
antigenic to each of the N robots). After the
antigenic robots and mines are known, the post-
affinity is calculated using
=
+
+
−
−
,
(2)
where:
• V corresponds to suppression from the
antibodies that represent robots competing for
the same mine (they may suppress the antigenic
ICAART 2012 - International Conference on Agents and Artificial Intelligence
8
robot if they have a higher fitness (lower
affinity) or are close in fitness to it).
• X corresponds to stimulation of antibodies that
represent robots competing for the same mine
(the antigenic robot may stimulate other robots
only if the other robots have a higher fitness
(lower affinity) than it).
• W corresponds to suppression from the
antibodies that represent mines competing for
use of the same robot (they may suppress the
antigenic mine if they have a higher fitness
(lower affinity) or are close in fitness to it).
• Y corresponds to stimulation of antibodies that
represent mines competing for use of the same
robot (the antigenic mines may stimulate other
mines only if the other mines have a higher
fitness (lower affinity) than it).
Equation (2) is similar to the original Farmer
equation but differs in two important respects. First,
concentrations of antibodies are not modelled, only
affinities, and second, there are two stimulation
terms and two suppression terms (rather than one of
each as in the original). This reflects the 2-
dimensional nature of the model used here, i.e.,
stimulation and suppression are considered between
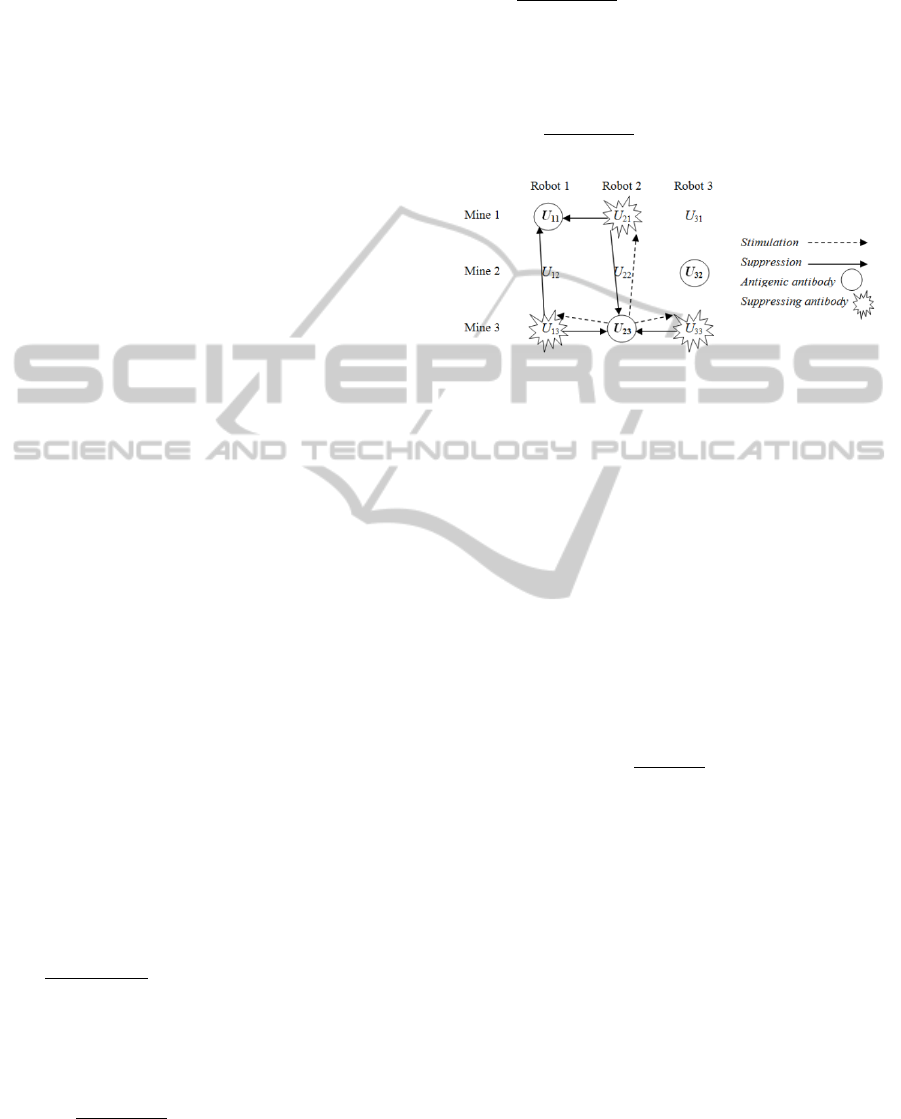
robots and also between mines. To illustrate, if the
affinities between mine-robot pairs were set out as a
matrix, for example with each row representing a
unique mine and each column representing a unique
robot, then stimulation and suppression are
measured both across the columns in the x-direction
and down the rows in the y-direction, see Figure 2,
which shows an example of 2-dimensional
stimulation and suppression for the 3-robot, 3-mine
case. Note also that stimulation terms are subtracted
from the pre-affinity and suppression terms are
added to it. This is because, in this case, the affinity
is based on the distance the robot has to travel, and
thus, a reduction is seen as an improvement.
In this model the total inter-robot suppression on
antibody γj is given by the sum of the suppressions V
imposed by antibodies kj (j = 1 to L, k = 1 to N)
where
=
−
∀ ≠ ⋀
−
<.
(3)
The inter-robot stimulation X on antibody ij is
imposed by antibody γj (i = 1 to N, j = 1 to L) and is
a single term given by
=
−
∀( ≠ ⋀
<
).
(4)
The total inter-mine suppression on antibody iγ is
given
by the sum of the suppressions W imposed by
antibodies ik (i = 1 to N, k = 1 to L) where
=
−
∀ ≠ ⋀
−
<.
(5)
The inter-mine stimulation Y on antibody ij is
imposed by antibody iγ (i = 1 to N, j = 1 to L) and is
a single term given by
=
−
∀( ≠ ⋀
<
).
(6)
Figure 2: An example of 2-dimensional stimulation and
suppression for a 3-robot, 3-mine case.
In the above equations,
is a scaling constant
that determines the overall level of stimulation and
suppression and is a constant that governs how
closely antibodies have to match in affinity to
become stimulated. After post-affinities have been
calculated the SBPA is implemented again to
allocate the new antigenic antibodies. The post-
affinities then become the new pre-affinities and
stimulation and suppression are calculated again.
The algorithm proceeds in this way until some
stopping criteria is met. The final, overall,
theoretical fitness F of the task-allocation solution is
determined as
=
10,000
∑
,
(7)
where
γj
is the distance between a final antigenic
robot and its allocated mine. Note that a different
measure of fitness, for example use of time taken t to
get to the mine (instead of d in the above equation)
should be used when attempting to demonstrate the
practical advantages of the ITTA, rather than the
theoretical. However, in the experiments described
here these measures are equivalent because of
assumptions 1 to 3.
The ITAA, as described above, is original in its
2-dimensional approach to stimulation and
suppression, its focus on affinities rather than
concentrations of antibodies, its novel suppression
and stimulation models, and its algorithmic
implementation, which results in the assignment of a
unique task to each robot, rather than the global
adoption of majority behaviours as in previous
AN IDIOTYPIC NETWORK APPROACH TO TASK ALLOCATION IN THE MULTI-ROBOT DOMAIN - Use of an
Artificial Immune System to Moderate the Greedy Solution
9
idiotypic research within the multi-robot domain.
Note that 1-dimensional models were trialled but
failed to guarantee converge to a solution. In
addition, a 2-dimensional model is a more accurate
reflection of an idiotypic system, where interactions
occur between all agents.
5 EXPERIMENTAL DETAILS
The ITTA was transcribed into MATLAB code and
was programmed to store the current fittest solution
after each iteration. The algorithm was stopped after
a maximum of 15 iterations had elapsed and the best
solution was accepted. In all cases, the initial
positions of the N robots and mines were generated
randomly on a square grid 30m by 30m in area, and
baseline comparisons were made for each problem
using the greedy (SBPA) algorithm (the solution
after the first iteration). Initially, the ITTA was
applied to 10,000 different mine diffusion problems
for N between 3 and 10 in order to determine
suitable values for parameters and
, i.e., the
above was repeated varying the parameter
between 10 and 1,000 (values of 10, 50, 100, 150,
250, 500, 750 and 1,000 were trialled), and varying
the parameter between 0.5m and 4m in steps of
0.5m. Once suitable values were found, the ITTA
was applied to a further 10,000 mine diffusion
problems for N ranging between 3 and 15, in order
to assess its performance against the baseline.
6 RESULTS
6.1 Parameter Selection
In all initial test cases
values of 10 and 50 proved
superior in performance to the others, with 10
tending to work better for smaller numbers of robots
(3 to 7) and 50 tending to work better for larger
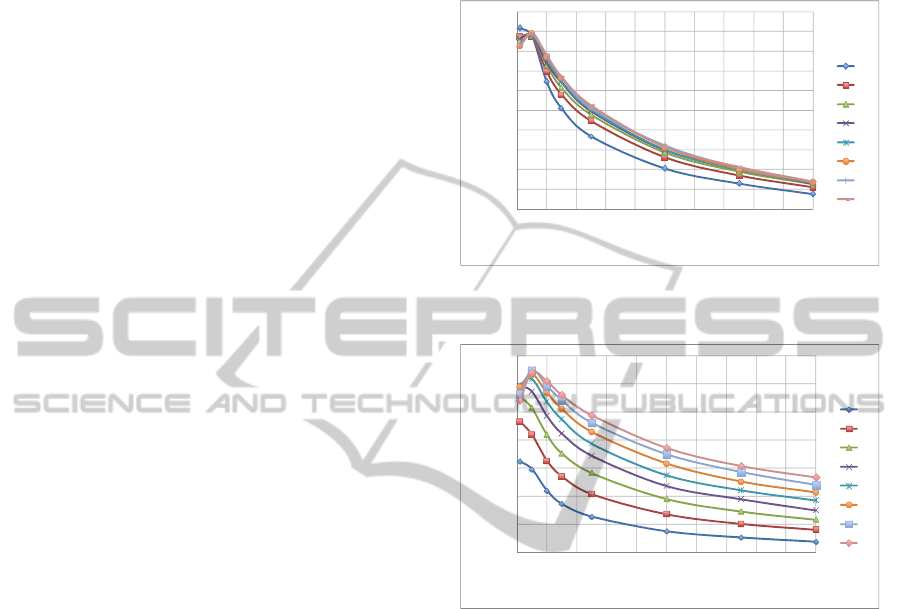
numbers (8 to 10). Figures 3a and 3b show how the
mean % improvement in fitness varies with
.
Figure 3a summaries the results for the different
values and Figure 3b does the same for the numbers
of robots N. Figure 3b also shows that mean %
improvement in fitness tends to increase steadily
with the number of robots; this is discussed more
fully in Section 6.2.
The value was more robust, demonstrating
much less variation in performance than
. This can
be seen in Figure 3a. Figures 4a and 4b also
summarise the preliminary results for ; the charts
show how mean % fitness improvement varies with
, with Figure 4a showing the results for each value
of N and Figure 4b showing the results for each
value of
.
Figure 3a: Variation of mean % improvement in fitness
with p
1
for different values.
Figure 3b: Variation of mean % improvement in fitness
with p
1
for different values of N.
Figure 4a also illustrates a clear trend for
increase in mean improvement in fitness as the
numbers of robots increases (see Section 6.2). In
general, there is a slight improvement as rises, but
the differences are much less pronounced than for
. Figure 4b highlights the poorer performance
when higher values of
are used. It shows that
values of either 10 or 50 are preferable and that
a
value of 50 has an almost constant performance
across the spectrum, whereas a
value of 10 tends
to work better for lower values of , between about
0.5 and 1.5.
As it showed a consistent performance for all
and worked well with higher numbers of robots, a
value of 50 was chosen for use in the performance
assessment, where N would rise to 15. Initially,
values of 3.0m and 4.0m were selected, based on the
preliminary results, but the best overall performance
was obtained when was set to 0.5m and
was set
to 50.
1.5
2
2.5
3
3.5
4
4.5
5
5.5
6
6.5
0 100 200 300 400 500 600 700 800 900 1000
Mean % Improvement in Fitness
p
1
0.5
1
1.5
2
2.5
3
3.5
4
ζ (m)
0.5
1.5
2.5
3.5
4.5
5.5
6.5
7.5
0 100 200 300 400 500 600 700 800 900 1000
Mean % Improvement in Fitness
p
1
3
4
5
6
7
8
9
10
N
ICAART 2012 - International Conference on Agents and Artificial Intelligence
10

Figure 4a: Variation of mean % improvement in fitness
with
for different values of N.
Figure 4b: Variation of mean % improvement in fitness
with
for different values of p
1
.
6.2 Performance Assessment
Table 1 summarises the performance of the ITTA
(using the assigned parameters,
= 50, = 0.5)
compared with the baseline greedy SBPA algorithm.
For all N there is an increase in the mean fitness of
the solution, which ranges from about 3.5% (N = 3)
to 7.2% (N = 9 and N = 11). Moreover, paired t-tests
conducted on the mean fitness values show that
ITTA fitness is significantly higher (at the 95%
level) than the baseline for all values of N, for all
results and also for the sub-set of improved cases.
Figure 5 shows that the mean improvement in fitness
starts off by increasing almost linearly with N but
gradually reaches a plateau at about N = 9. This
may be explained as follows; the likelihood that the
initial, greedy solution may already be the optimal
one is higher for smaller N, and so there is less room
for improvement. This explanation is validated by
examination of the % of improved cases, which also
increases steadily with N up until reaching a plateau
at about N = 10, see Figure 6. In addition, when the
improved cases are examined in isolation, as
expected, the % improvement is greater for all N, but
this
is more pronounced for smaller N, for example,
Table 1: Performance summary for ITTA and baseline.
the difference is 8.25% for N = 3, but only about
1.30% at plateau values of N, see Figure 5.
For all N the maximum % improvement in
solution is considerably higher than the mean, for
example, for N = 3 it is about 76% and for N = 12 it
is about 46%. This variable tends to oscillate
locally, but has a general downward trend with
increased N, see Figure 6. For all results, the mean
number of iterations ranges from about 2.0 for N = 3
to about 6.5 for plateau values of N, see Figure 7,
which is intuitive given the earlier explanation for
plateau behaviour. For improved cases only this
variable is much more consistent, tending to about
7.0 iterations.
Figure 5: Variation of mean % improvement in fitness
with N.
The above results suggest that the ITTA is able
to make significant improvements over greedy
strategies, and that, as the number of robots
increases, an improvement in the solution is more
likely. For the particular parameters used here there
is approximately an 85% chance of generating a
1.5
2.5
3.5
4.5
5.5
0.511.522.533.54
Mean % Improvement in Fitness
ζ (m)
3
4
5
6
7
8
9
10
N
1.5
2.5
3.5
4.5
5.5
6.5
0.511.522.533.54
Mean % Improvement in Fitness
ζ (m)
10
50
100
150
250
500
750
1000
p
1
3
4
5
6
7
8
9
10
11
12
3456789101112131415
Mean % Improvement in Fitness
N
Improved cases only All results
AN IDIOTYPIC NETWORK APPROACH TO TASK ALLOCATION IN THE MULTI-ROBOT DOMAIN - Use of an
Artificial Immune System to Moderate the Greedy Solution
11
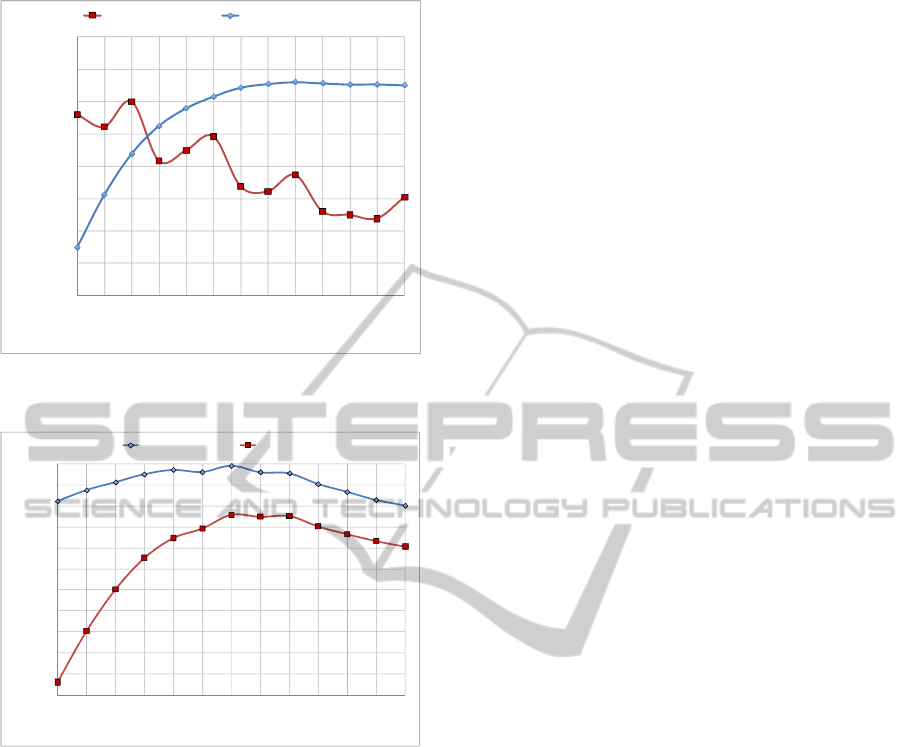
Figure 6: Variation of max % improvement in fitness and
% of improved cases with N.
Figure 7: Variation of mean iterations with N.
better solution for N greater than 8. For N greater
than 6, a mean increase in fitness of about 7% is
expected, although individual increases of up to
about 70% are possible. The ITTA has also proved
to be a fast algorithm as the mean number of
iterations for convergence is always below eight.
In addition, a further set of experiments that
varied
between 50 and 150 across the suppression
and stimulation equations (3), (4), (5) and (6) has
also been conducted for N = 4. The use of
= 50 in
all equations except (3) (which used 90 instead)
increased the overall performance by a further 0.7%.
These results demonstrate the potential of the
ITTA method and show that it is a good candidate
for further investigation involving more complex
problems (as described fully in Section 3), real-
world implementations and more rigorous parameter
tuning. Preliminary investigations have already
shown that the method is easily adapted to cases
where L > N and N > L. Where there are more robots
than mines (N > L) robots are simply marked as
redundant when the SBPA part of the algorithm does
not allocate them to a mine. In addition, ITTA
consistently outperforms SBPA and, as N increases
for a fixed number of L, performance improves.
Conversely, when L > N absolute performance
drops, with the algorithm having to run repeatedly as
robots change position, but ITTA still performs
better than the greedy algorithm. Thus, relatively
speaking, there is no noticeable drop in ITTA's
performance when L > N.
7 FUTURE WORK
Future work will aim to develop an optimum
stopping criteria and to test the algorithm in more
complex scenarios, where online and time-extended
assignments are required, there are heterogeneous
tasks and robots, multi-task robots and multi-robot
tasks. Real-world implementations that require
additional features such as reactive obstacle
avoidance modules will also be carried out in an
outdoor environment. Further work also needs to be
done to compare performance of the ITTA with
state-of-the-art task allocation methods (for example
market-based approaches) and linear optimization
techniques such as Mixed Integer Linear
Programming (MILP).
Note that in real-life implementations the
algorithm would need to run independently on each
robot in order to constitute a truly decentralized
system. The robots would also need to communicate
reliably in order to transmit their locations to one
another, and there would need to be a level of
assurance that each robot was receiving all the
available information and compiling the same
solution to the problem. Maintaining and sharing an
accurate intelligence picture within an ad-hoc
network has been the subject of a research program
within BAE Systems Advanced Technology Centre
(ATC), and the outputs have already produced a
prototype data sharing framework. Future work will
thus aim to integrate the ideas presented in this paper
with the outputs of the data sharing programme in
order to demonstrate decentralized multi-robot task-
allocation in a real-world environment. In addition,
the ATC has also been working on Consensus-Based
Bundle Algorithms (CBBA) and Max-Sum task
allocation mechanisms (Mathews et al. (2010)), so
work will be undertaken to assess the feasibility of
integrating the ITTA approach with those methods
(see also Stranders et al. (2009)).
20.00
30.00
40.00
50.00
60.00
70.00
80.00
90.00
100.00
3456789101112131415
Percentage
N
Max % improvement % of improved cases
2.00
2.50
3.00
3.50
4.00
4.50
5.00
5.50
6.00
6.50
7.00
7.50
3 4 5 6 7 8 9 101112131415
Iterations
N
Improved cases only All results
ICAART 2012 - International Conference on Agents and Artificial Intelligence
12

8 CONCLUSIONS
This paper has described an idiotypic AIS algorithm
(ITAA) for solving task allocation problems in the
multi-robot domain. The algorithm is novel since
other idiotypic approaches have only been
applicable to problems where many robots are
required to perform one task at a time using
swarming behaviours; in contrast ITTA is suited to
problems that require members of a multi-robot team
to act individually so that different tasks can be
solved simultaneously. The algorithm is also original
in its implementation of the Farmer equation, which
ignores concentrations of antibodies and uses novel,
2-dimensional models for stimulation and
suppression of the antibody affinities.
A series of initial tests have been carried out on
the algorithm using simulated mine diffusion
problems in MATLAB. These tests have helped to
establish suitable parameter values for the
stimulation and suppression terms and have
provided statistical evidence that the ITTA is
capable of out-performing the greedy Sequential
Best-Pair Assignment (SBPA) algorithm in about
85% of cases for numbers of robots N exceeding 8.
For smaller N the likelihood of outperforming the
greedy solution rises almost linearly as N increases.
The ITTA has also shown fast convergence to a
solution; for N of 8 and above the mean number of
iterations for arrival at the best solution is about 5,
i.e., the solution can be produced almost
instantaneously.
REFERENCES
Atay, N., Bayazit, B., 2007. Emergent task allocation for
mobile robots. In Proceedings of the Robotics: Science
and Systems Conference (RSS’07), Atlanta, GA, USA.
Barra, A., Agliari, E., 2007. Stochastic dynamics for
idiotypic immune networks. Physica A 389: pp. 5903-
5911.
Burnet, F.M., 1959. The clonal selection theory of
acquired immunity, Cambridge University Press,
Cambridge, U.K.
Dias M. B., Stentz, A., 2002. Opportunistic optimization
for market-based multi-robot control. In Proceedings
of the IEEE/RJS International Conference on
Intelligent Robots and Systems (IROS), Lausanne,
Switzerland: pp. 2714-2720
Dioubate, M., Guanzheng, T., Toure-Mohamed, L., 2008.
An artificial immune system based multi-agent model
and its application to robot cooperation problem. In
Proceedings of the 7th World Congress on Intelligent
Control and Automation, Chongging, China: pp.
3033-3039.
Farmer, J. D., Packard, N. H., Perelson, A. S., 1986. The
immune system, adaptation, and machine learning.
Physica, D, 2(1–3): pp. 187–204.
Fukuda T., Nakagawa, S., Kawauchi, Y., Buss, M., 1988.
Self-organizing robots based on cell structures -
CEBOT. In Proceedings of the IEEE/ RSJ
International Conference on Intelligent Robots and
Systems (IROS), Victoria, British Columbia, Canada:
pp. 145-150.
Gerkey, B. P., Mataric, M. J., 2002. Sold! Auction
methods for multi-robot coordination, IEEE
Transactions on Robotics and Automation 18(5): pp.
758-768.
Gerkey, B. P., Mataric, M. J., 2004. A formal analysis and
taxonomy of task allocation in multi-robot systems.
International Journal of Robotics Research 23(9): pp.
939-954.
Jerne, N. K., 1974. Towards a network theory of the
immune system. Ann. Immunol. (Inst Pasteur),
125C(1/2): pp. 373–389.
Jun, J-H., Lee, D-W., Sim, K-B.,1999. Realization of
cooperative strategies and swarm behavior in
distributed autonomous robotic systems using artificial
immune system. In Proceedings of the 1999 IEEE
International Conference of Man and Cybernetics 6:
pp. 614-619. IEEE Press, New York.
Lee, D-W., Sim, K-B., 1997. Artificial Immune Network-
based Cooperative Control in Collective Autonomous
Mobile Robots. In Proceedings of the IEEE
International Workshop on Robot and Human
Communication: pp. 58-63.
Li, J., Xu, H., Wang, S., Bai, L., 2007. An immunology-
based cooperation approach for autonomous robots. In
Proceedings of the 2007 International Conference on
Intelligent Systems and Knowledge Engineering (ISKE
2007), 4.
Liu, W., Winfield, A. F. T., Sa1, J., Chen, J., Dou, L.,
2010. Towards energy optimization: Emergent task
allocation in a swarm of foraging robots. Adaptive
Behavior 15 (3): pp. 289-305
Luh G. C., Liu, W.W., 2004. Reactive immune network
based mobile robot navigation. In Proceedings of the
3rd International Conference on Artificial Immune
Systems (ICARIS) 3239: pp. 119–132.
Mathews, G., Waldock, A., Paraskevaides, M., 2010.
Toward a decentralised sensor management system for
target acquisition and track. In Proceedings of the 5th
SEAS DTC Technical Conference: pp. C2.
Mitsumoto, N., Fukuda, T., Shimojima, K., Ogawa, A.,
1995. Micro autonomous robotic system and
biologically inspired immune swarm strategy as a
multi agent robotic system. In Proceedings of the 1995
IEEE International Conference on Robotics and
Automation: pp. 2187-2192.
Nanjanath, M., Gini, M., 2010. Repeated auctions for
robust task execution by a robot team. Journal
Robotics and Autonomous Systems 58(7): pp. 900-909,
North-Holland Publishing Co. Amsterdam, The
Netherlands.
AN IDIOTYPIC NETWORK APPROACH TO TASK ALLOCATION IN THE MULTI-ROBOT DOMAIN - Use of an
Artificial Immune System to Moderate the Greedy Solution
13

Oliver, G., Guerrero, J., 2011. Auction and swarm multi-
robot task allocation algorithms in real time scenarios.
In Multi-Robot Systems, Trends and Development,
Toshiyuki Yasuda (Ed.), ISBN :978-953-307-425-2:
pp. 437-456, InTech.
Razali, S., Meng, Q., Yang, S-H., 2009. Multi-robot
cooperation using immune network with memory. In
Proceedings of the 2009 IEEE International
Conference on Control and Automation, Christchurch,
New Zealand: pp. 145-150.
Razali, S., Meng, Q., Yang, S-H., 2010. A refined immune
systems inspired model for multi-robot shepherding.
In Proceedings of the 2010 Second World Congress on
Nature and Biologically Inspired Computing,
Kitakyushu, Fukouka, Japan: pp. 473-478.
Sathyanath, S., Sahin, F., 2002. AISIMAM—An AIS
based intelligent multi-agent model and its application
to a mine detection problem. In Proceedings of the 1st
International Conference on Artificial Immune
Systems (ICARIS).
Shehory, O., Kraus, S., 1998. Methods for task allocation
via agent coalition formation, Artificial Intelligence
101(2): pp. 165-200.
Stranders, R., Farinelli, A., Rogers, A., Jennings, N. R.,
2009. Decentralised coordination of mobile sensors
using the max-sum algorithm. In Proceedings of the of
the Twenty-First International Joint Conference on
Artificial Intelligence (IJCAI-09): pp. 299-304.
Sun, S-J, Lee, D-W., Sim, K-B., 2001. Artificial immune-
based swarm behaviors of distributed autonomous
robotic systems. In Proceedings of the 2001 IEEE
International Conference on Robotics and Automation
(ICRA 2001), 4: pp. 3993-3998.
US Department of Defence, 2005. Unmanned Aircraft
Systems Roadmap 2005-2030.
US Department of Defence, 2009. Unmanned Systems
Integrated Roadmap FY2009-2034.
Vargas, P. A., de Castro, L. N., Michelan, R., 2003. An
immune learning classifier network for autonomous
navigation. In Proceedings of the 2nd International
Conference on Artificial Immune Systems (ICARIS)
2787: pp. 69–80.
Watanabe, Y., Ishiguro, A., Shirai, Y., Uchikawa, Y.,
1998. Emergent construction of behavior arbitration
mechanism based on the immune system. In
Proceedings of IEEE ICEC: pp. 481–486.
Whitbrook, A. M., Aickelin, U., Garibaldi, J. M., 2007.
Idiotypic Immune Networks in Mobile-Robot Control.
In IEEE Transactions on Systems, Man, And
Cybernetics—Part B: Cybernetics, 37(6): pp. 1581-
1598.
ICAART 2012 - International Conference on Agents and Artificial Intelligence
14
