
TOWARDS AN IMMUNE-INSPIRED TEMPORAL ANOMALY
DETECTION ALGORITHM BASED ON TUNABLE ACTIVATION
THRESHOLDS
M´ario Antunes
1
, Manuel Correia
2
and Jorge Carneiro
3
1
School of Technology and Management, Polytechnic Institute of Leiria, Morro do Lena-Alto do Vieiro, Leiria, Portugal
2
Department of Computer Science, University of Porto, Rua do Campo Alegre, 1021-1055, Porto, Portugal
3
Instituto Gulbenkian de Ciˆencia, Oeiras, Portugal
Keywords:
Artificial immune system, Anomaly detection, Tunable activation threshold, T-cell simulation and modelling,
Pattern recognition.
Abstract:
The detection of anomalies in computer environments, like network intrusion detection, computer virus or
spam classification, is usually based on some form of pattern search on a database of “signatures” for known
anomalies. Although very successful and widely deployed, these approaches are only able to cope with anoma-
lous events that have already been seen. To cope with these weaknesses, the “behaviour” based systems has
been deployed. Although conceptually more appealing, they have still an impractical high rate of false alarms.
The vertebrate Immune System is an emergent and appealing metaphor for new ideas on anomaly detection,
being already adopted some algorithms and theoretical theories in particular fields, such as network intrusion
detection. In this paper we present a temporal anomaly detection architecture based on the Grossman’s Tun-
able Activation Threshold (TAT) hypothesis. The basic idea is that the repertoire of immune cells is constantly
tuned according to the cells temporal interactions with the environment and yet retains responsiveness to an
open-ended set of abnormal events. We describe some preliminary work on the development of an anomaly
detection algorithm derived from TAT and present the results obtained thus far using some synthetic data-sets.
1 INTRODUCTION
The vertebrate Immune System (IS) (Sompayrac,
2008) inspired the deployment of Artificial Immune
Systems (AIS) ( de Castro and Timmis, 2002) and has
already been successfully used as a promising source
of inspiration for new ideas on anomaly detection
(Kim et al., 2007). The IS is an extremely complex
distributed system whose main function is to actively
protect the body from the intrusion of pathogens. It
is composed by two main layers of defense: innate
and adaptive. The innate part only recognizes specific
known intruders by their “signatures”, and its behav-
ior is similar in all individuals of the same species.
In contrast, the adaptive part is in a sense unique to
each individual and is able to “learn” throughout time
to recognize new forms of intrusive pathogens, thus
providing a much more specific and adaptive form of
recognition of pathogens.
The IS is supported by a complex set of cells. The
Antigen Presenting Cell (APC) digests and converts
pathogens into small peptides which are then pre-
sented to T-cells. These cells have specific receptors
that can bind with a certain degree of affinity to the
peptides present on the surface of each APC and thus
become activated.
Anomaly detection can be seen as a technique that
produces a model for identifying cases that in some
way deviate from a “learned” normal behavior. Deci-
sions are based on a profile of normal behaviour and
an anomaly is any particular case instance that is an
outlier under this characterization. Current anomaly
detection systems are mainly based on statistics, data-
mining, data fusion and bio-inspired approaches, like
neural networks.
Interestingly, the problem of creating a system ca-
pable of monitoringa normally changingenvironment
and yet retaining the capacity to detect open-ended
anomalies has been developed by natural selection
during the evolution of the vertebrate IS. This sys-
357
Antunes M., Correia M. and Carneiro J. (2009).
TOWARDS AN IMMUNE-INSPIRED TEMPORAL ANOMALY DETECTION ALGORITHM BASED ON TUNABLE ACTIVATION THRESHOLDS.
In Proceedings of the International Conference on Bio-inspired Systems and Signal Processing, pages 357-362
DOI: 10.5220/0001553303570362
Copyright
c
SciTePress

tem is capable of discriminate and engage in very dif-
ferent ways both normal body components and very
similar but foreign (abnormal) chemical structures
present in microorganisms. The IS is also able to learn
and memorise the first encounter it has with these in-
truders, and can make effective use of this acquired
knowledge to better deal with them on a future en-
counter. Perhaps even more relevant for the designing
of an effective anomaly detection system, is the now
well accepted fact that the IS learns the body compo-
sition during embryo life and adapts to physiological
changes as the individual matures and ages (notable
examples being hormones during sexual maturation
or metamorphoses in some vertebrates).
Negative Selection (NS) (Forrest et al., 1994) and
Danger Theory (DT) (Greensmith et al., 2006) were
the immune theories most used on the development
of IS-based anomaly detection systems (Kim et al.,
2007). In this paper we explore a different view of
the immune system to present the first developments
of a new anomaly detection algorithm based on the
Tunable Activation Threshold (TAT) hypotheses put
forward by Grossman and colleagues (Grossman and
Paul, 1992). In TAT it is assumed that lymphocytes
have tunable activation thresholds whose value re-
flects the recent history of signaling they have been
receiving from the environment. Potentially autoim-
mune lymphocytes, which are continuously exposed
to body antigens raise their activation threshold, and
become unresponsive or anergic. In contrast, lympho-
cytes that are not auto-reactive but recognise microor-
ganism structures have low activation thresholds and
are thus fully responsive upon infection.
This paper is organized as follows: in section 2 we
explain in some detail the TAT concept and the model
dynamics we have used for T-cells. In section 3 we
describe the system architecture, its main components
and features, as well as a methodology we have used
for the generation of synthetic data-sets we are using
to evaluate the system in a controlled way. In section
4 we present the results obtained with the experiences
we have done with the artificially generated generic
data-sets. In section 5 we discuss the results obtained,
draw some conclusions and delineate guidelines for
future research.
2 TAT AS A MODEL FOR
TEMPORAL ANOMALY
DETECTION
The Tunable Activation Threshold (TAT) (Grossman
and Paul, 1992) hypothesizes that T-cell activation de-
pends on a threshold that is adjusted dynamically to
the integrated history of signals received via the T-
cell Receptor (TCR). Every interaction between the
TCR and its ligands, the antigenic MHC/peptide com-
plexes presented by the APC, results in intracellular
competition between ”excitation” and ”de-excitation”
signaling pathways, causing the T-cell to adapt to
the stimulus by increasing or decreasing its activation
threshold. Therefore, T-cells with different antigen-
specificity will have different activation thresholds as
they are exposed to different stimuli. Furthermore,
Grossman and colleagues also postulated that T-cells
that are tuned to be unresponsive by chronic exposure
to antigen could inhibit the activation of responsive T-
cells in their neighborhood in physical and antigenic
spaces. This implies that an immune response will
not depend on response of an individual T-cell, but
depends on the ensemble of T-cells engaged and on
their current activation thresholds, which in turn re-
flects the T-cell’s individual history.
We have adopted a minimal mathematical model
of TAT for T-cells (Carneiro et al., 2005). Briefly,
T-cell activation is controlled by two enzymes that re-
spond to antigenic signals delivered by the APC: Ki-
nase (K) and Phosphatase (P). Antigenic signals lead
to a linear increase of both K and P activities until
they reach a plateau that is proportional to the inten-
sity of the stimulus.
0
100
200
300
400
500
600
0 100 200 300 400 500 600
Signal activity
t
TESTING - T-cell: abcd
K
P
S
Figure 1: The T-cell receives a variable signal and adjusts
the K and P levels.
For the same signal S, K increases faster than P,
but if the signal persists P will eventually reach a
higher plateau. Similarly, on signaling absence, K re-
turns to the basal level at a faster rate than P. It is
further assumed that T-cell activation is a switch-type
response that requires that K supersedes P, at least
transiently. Under these conditions, those T-cells that
receive continuous or sufficiently frequent antigenic
signals from APCs become unresponsive and those
that rarely see their antigen remain sensitive (Carneiro
et al., 2005) (illustrated in Figure 1).
BIOSIGNALS 2009 - International Conference on Bio-inspired Systems and Signal Processing
358

3 GENERAL ARCHITECTURE
In this section we present the TAT-based architecture
we developed for anomaly detection. We describe a
TAT-based AIS and its major metaphorical IS coun-
terparts for anomaly detection.
Figure 2: Building blocks of the TAT-based AIS.
Figure 2 depicts the general architecture of TAT-
based AIS. The core is a simulation of the dynamics
of an artificial T-cell repertoire. Each artificial T-cell,
heretofore denoted by TCELL, is an object that re-
ceives signals from the environment, compares these
signals to a string representation of its unique speci-
ficity, and adjusts its response threshold by tuning the
values of its K and P variables. At some given point in
time, a ”committee” of activated TCELLs may raise
an alert depending on the values each cell has for
these variables.
3.1 Generating Artificial Data-Sets
We have worked with two different data-sets: one is
used for training and the other is used for testing.
The training data-set is further split in two parts, re-
specting the temporal order of the events recollec-
tion. The first part is used for training calibration
in which the T-cell simulator is run, neglecting any
alerts. The second part is composed by normal and
abnormal APCs, where the abnormal APCs contain
peptides not present on the first part of the training
data-set. The aim of the training phase is to build and
tune the TCELL repertoire and adapt the values of
K and P to the environment. The second part of the
training data-set is used to validate the system initial
calibration. This is done by evaluating how well the
newly constructed cell repertoire copes with the APCs
present in the second part of the training data-set.
Each APC is a container composed by a set of
string PEPTIDEs separated by a white space and a
classification tag. On the experiments described in
this paper, the data-sets have been synthetically gen-
erated by having the PEPTIDEs for each APC taken
from a group of pre-arranged “string” sets, as follow:
a set of normal PEPTIDEs (strings) that appear reg-
ularly on both training and testing data-sets; a set rep-
resenting sporadic patterns that appear in training, but
are also considered normal; two sets of patterns corre-
sponding to anomalies in training and testing respec-
tively; finally, a set of new patterns for testing that
were unseen during training, but are still the result of
normal activity.
We have generated (Section 4) artificial data-sets
that meet the following conditions: the training data-
set has 5000 APCs (3750 for calibration and 1250 for
validation); the testing data-set has 7500 APCs. The
APCs have a maximum number of 1000 PEPTIDEs,
generated randomly from the sets described above.
The APCs with anomalies are different from those
generated for the testing and for the training data-sets.
3.2 TCELL Repertoire Dynamics
The AIS contains a variable list of TCELLs that are
dynamically created and deleted. Each TCELL has
a unique string, which defines its specificity, and is
analogous to the TCR of the natural T-cell. It also
stores two variables, K and P, that are adjusted as a
function of the input signal S receivedfrom each APC,
as described in next section. A TCELL is created and
added to the repertoire whenever a PEPTIDE in any
of the currently queued APC does not find a suffi-
ciently similar match in the available repertoire. In
this newly created TCELL the string is set to be iden-
tical to the unmatched PEPTIDE. The K and P are
initially set to the basal values (K
0
and P
0
, respec-
tively) and updated with the stimulus represented by
the PEPTIDE.
A TCELL is removed from the repertoire when-
ever the K and P dynamics bring them back to these
basal values. In practice, this algorithm of creation
and removal of TCELLs, uses implicitly a potential
infinite repertoire of TCELLs with K and P in basal
values, but we only use the processing and memory
resources upon demand, keeping the actual repertoire
size contained.
3.3 TCELL K and P Dynamics
The TAT model we implemented is a piece-wise lin-
ear approximation to the differential equations model
described in (Carneiro et al., 2005), and can be de-
scribed as follows:
1. T-cells are born with basal values of K and P, re-
spectively K
0
= S
0
∗ K
max
and P
0
= S
0
∗ P
max
. S
0
corresponds to the initial value for the signal re-
ceived by T-cells.
2. The values of K and P are adjusted dynamically as
a function of the signal S and tend towards the val-
ues K
0
+ S∗ K
max
and P
0
+ S∗ P
max
, respectively.
TOWARDS AN IMMUNE-INSPIRED TEMPORAL ANOMALY DETECTION ALGORITHM BASED ON TUNABLE
ACTIVATION THRESHOLDS
359

3. The input signal S sent by each PEPTIDE to the
TCELLs in the repertoire is calculated by:
S = C× Af finity(TCELL, PEPTIDE)
where C is the number of occurrences of the
PEPTIDE in the APC and Af finity is the per-
centage of equal characters in the same positions,
for all the TCELL and PEPTIDE strings.
4. If the values of K and P are lower (higher) than
their maximum values, they increase (decrease)
linearly with constant derivatives φK and φP, re-
spectively, until the corresponding plateau values
are reached (step 2).
5. The TCELL is transiently activated when K ¿ P;
otherwise it is said to be unresponsive.
6. To ensure that a TCELL receiving a constant sig-
nal eventually becomes unresponsive (Figure 1)
we impose P
0
+ S∗ P
max
> K
0
+ S∗ K
max
.
7. To ensure that, in any TCELL, the condition K ¿ P
can be potentially reached at least transiently, we
impose that φK > φP.
8. The time duration of each APC is measured in
units of APC
duration
(∆t).
Algorithm 1 shows the corresponding pseudo-
code for the updating of TCELL variables.
Algorithm 1 Update TCELL parameters.
1: if ((S+ S
0
) ∗ K
max
) > K then
2: K ← MIN((S+ S
0
) ∗ K
max
, K+ = φK ∗ ∆t)
3: else
4: K ← MAX((S+ S
0
) ∗ K
max
, K− = φK ∗ ∆t)
5: end if
6: if ((S+ S
0
) ∗ P
max
) > P then
7: P ← MIN((S+ S
0
) ∗ P
max
, P+ = φP∗ ∆t)
8: else
9: P ← MAX((S+ S
0
) ∗ P
max
, P− = φP∗ ∆t)
10: end if
3.4 The Immune Response in the AIS
In the AIS, the APCs currently queued are processed
sequentially, reflecting the temporal order of events,
and are classified according to the activation state of
the TCELLs that match its PEPTIDEs. A TCELL
is considered to match a PEPTIDE if their pairwise
Af finity is greater than a predefined value α. The
classification of the APC is decided based on the com-
mittee of all TCELLs matching its PEPTIDEs. We
first compute the fraction of activated TCELLs per
PEPTIDE, and count the number of PEPTIDEs in
which this fraction is greater than a critical value τ.
If the number of such ’abnormal’ peptides relative to
the number of PEPTIDEs in the APC is higher than a
predefined parameter ψ, then an alert is raised against
the APC.
3.5 Adjusting the System
The parameters controlling the natural IS have been
slowly refined by millions of years of selection of
ancestrals who managed to defend themselves from
pathogens, and yet avoided autoimmunity. Similarly,
we set the run-time parameters of the TAT algorithm
by running a non-linear meta-heuristic simplex op-
timizer (Pedroso, 2007). The mission of this opti-
mizer is to make sure that the TAT-based AIS clas-
sifies properly the APCs generated over an appropri-
ate training data-set, tuning automatically the TCELL
repertoire to the environment. The optimizer uses
only the APCs generated during the training valida-
tion to compare the classification made by the AIS
algorithm with the classification tag. The optimizer
runs the AIS algorithm repeatedly over the bipartite
data-set until it finds the parameter regime in which
the TAT algorithm tunes the repertoire and is able to
raise alerts on the APCs containing artificial anoma-
lies, with a minimal number of false alerts on other
APCs.
The introduction of at least one artificial anomaly
in the training set for parameters validation is abso-
lutely necessary to constrain the tuning dynamics to
meaningful parameter regimes. In our experiments
we observed that if we would only use normal events
in training data-set and require minimization of false
alarm rates during the training validation, the sim-
plest solutions returned by the optimizer are param-
eter regimes in which tuning of K and P is so strong
that no TCELL could ever be activated. This allowed
us to better guide the optimizer in finding a set of
parameters that, not only minimizes the rate of false
alarms, but can also achieve a low rate for false neg-
atives. In addition we introduced a few additional
heuristic constraints:
0 <
K
0
P
0
< 1 φK > φP 0 <
K
max
P
max
< 1
The values for K
max
, φK, K
0
and S
0
have also been
fixed at 10, 15, 100 and 10 respectively. Also, the
values of α, τ and ψ were also optimised.
4 TESTING THE TAT-BASED AIS
Our working hypothesis is that a TAT-based system
with the architecture just described could be opti-
mised in such a way that its TCELL repertoire would
BIOSIGNALS 2009 - International Conference on Bio-inspired Systems and Signal Processing
360
be tuned to the individual characteristics of a real
environment and yet be able to raise alerts against
anomalous activity. In that sense, the main achieve-
ment is the development of an implementation of the
framework we have previously presented. We have
already obtained some results with our implementa-
tion, which are described and discussed in the follow-
ing sections.
4.1 Experimental Protocol
We conducted two sets of five experiments each, with
different data-sets. In the first set, each anomaly is
extended between two contiguous APCs. Our aim is
to detect the region (one or both APCs) that overlaps
with the anomaly. In the second experiment we in-
tend to decide whether TAT could also be used as an
APC classifier. To be able to determine this, in the
second data-set, anomalies appear isolated within a
single APC. We fixed the maximum number of APCs
with anomalies in 200 in the training data-set. In the
testing data-set we started with 149 anomalies and in-
crease this value in the subsequent experiments. Ta-
ble 1 describe the optimised parameters sets for each
of the data-sets, using the fixed parameters described
in subsection 3.5. The upper 5 rows correspond to
the “detection” of contiguous APCs and the lower 5
rows correspond to the best parameters that correctly
classified the APCs with anomalies.
4.2 Results
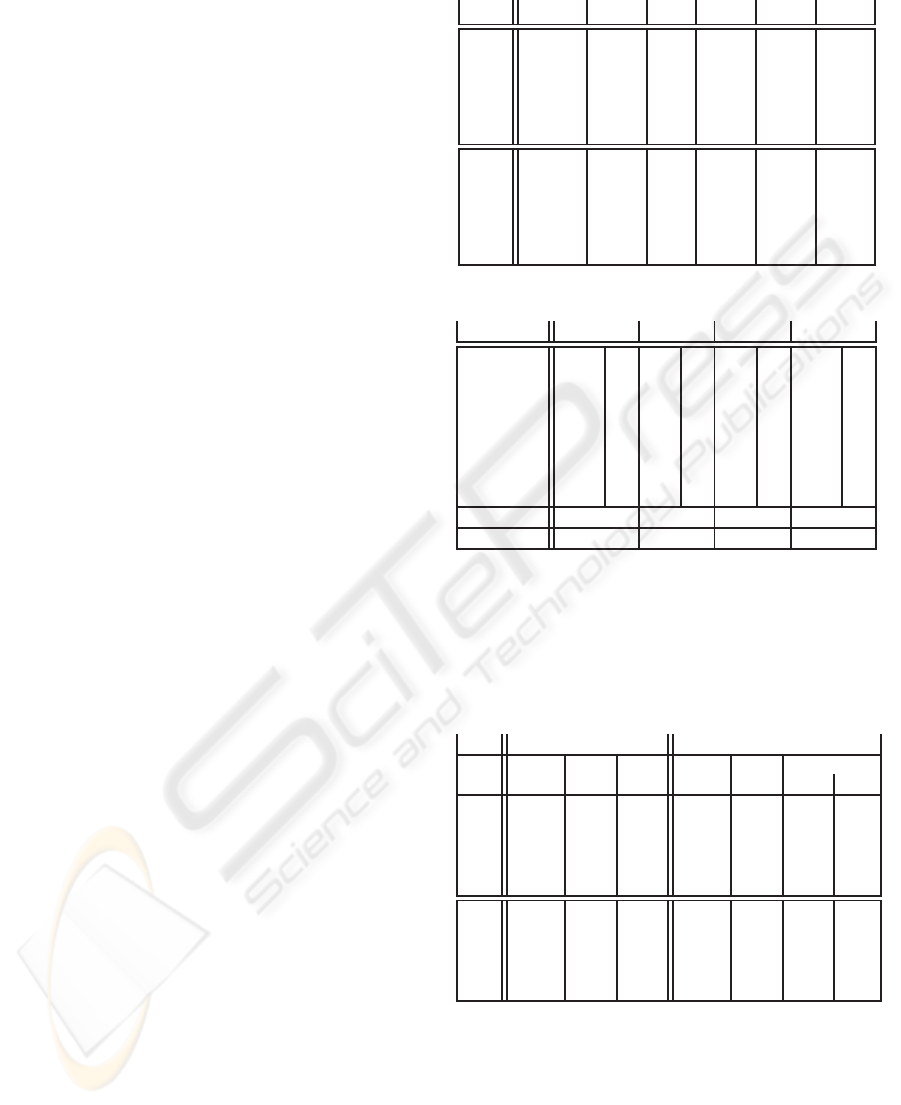
In order to evaluate the characteristics of TAT for de-
tection, let us assume that C is the percentage of rare
peptides in the APC. In table 2 we consider that the
parameter ψ was optimised to the value of 6%, which
means that if the ratio of abnormal peptides in the
APC is above this value, then the APC is considered
abnormal and an alarm should be raised. For each
APC we show the concentration of each PEPTIDE
and the decision made by the AIS. In the bottom we
present the classification of each APC.
In this example, both APCs 2 and 3 have ab-
normal PEPTIDEs and thus should both be classi-
fied as “abnormal“. Nevertheless, since the abnormal
PEPTIDEs doesn’tmatch any T-cell in the repertoire,
new ones are created with the initial values of K and
P. According to TAT dynamics (Section 3.3), the sig-
nal S (peptide concentration times the affinity) sent by
the APC should be such that K become higher that P.
Thus, the region where the anomaly took place com-
prises the APCs 2 and 3 and the AIS raised an alarm in
the APC 3. The two APCs did not have been correctly
classified, but the region where the anomaly happened
Table 1: TAT optimised parameter set.
Run P
0
P
max
φ
P
α τ ψ
1 188.4 18.8 2.2 11.1 6.4 88.3
2 127.4 12.7 9.3 38.1 10.1 13.8
3 138.9 13.1 7.9 6.9 2.0 78.3
4 125.1 12.5 9.4 72.9 15.0 54.4
5 144.5 14.4 5.6 57.5 7.3 56.6
1 131.3 13.1 5.8 16.1 8.5 67.9
2 137.1 13.7 3.2 4.32 2.71 77.8
3 127.6 12.7 2.6 80.1 11.8 83.3
4 124.7 12.4 7.3 77.1 10.2 48.7
5 151.4 15.1 4.1 10.4 6.83 55.9
Table 2: Artificial immune detection. ψ = 6%.
PEPTIDE APC
1
APC
2
APC
3
APC
4
abcde 84 N 33 N 37 N 107 N
fghij 97 N 53 N 45 N 99 N
klmno 101 N 41 N 36 N 79 N
pqrst 89 N 40 N 39 N 99 N
uvwxz 97 N 53 N 42 N 101 N
PQRST - - 43 N 40 Y - -
UVWXZ - - 36 N 29 Y - -
OOOOZ - - 43 N 29 Y - -
C(%) 0 0 32% 0
Decision N N Y N
was correctly detected.
With the resulting optimised parameters sets for
the experiences (Table 1), and using the ”committee”
classification algorithm (Algorithm 1), we obtained
the results described in Table 3.
Table 3: Results obtained during the experiments.
Training Testing
Run APC TP FP APC
TP FP
qty qty %
1 192 121 15 149 75 14 0.1
2 198 129 20 199 104 66 0.8
3 195 122 17 248 131 34 0.4
4 194 127 18 299 163 83 1.1
5 198 126 23 347 184 31 0.4
1 192 192 18 149 148 21 0.2
2 198 198 12 199 198 46 0.6
3 195 195 159 248 248 563 7.5
4 194 193 92 299 299 417 5.5
5 198 195 27 347 347 35 0.4
Each row represents the best results obtained with
the parameters presented on Table 1, for each ex-
periment. The upper 5 rows correspond to the ex-
periments with two contiguously generated anomaly
APCs and the lower 5 with isolated anomaly APCs.
In the first experiments, the TAT algorithm could de-
tect at least one of the APC carrying the anomaly,
which appeared contiguously in the testing data-set.
As can be seen, the true positives corresponds to more
TOWARDS AN IMMUNE-INSPIRED TEMPORAL ANOMALY DETECTION ALGORITHM BASED ON TUNABLE
ACTIVATION THRESHOLDS
361

than half of the total APCs, which means that all
the anomalies was detected and, in some cases, both
APCs of an anomalies raised an alert. We was also
succeeded in the last 5 experiments, whose goal was
to evaluate the use of TAT for classification. In the
testing phase we have a high rate of true positive and
a relatively low rate of false alarms. The higher level
of false alarms was 7.5% in the third experiment.
5 DISCUSSION
We have described an algorithm for anomaly detec-
tion based on the TAT theoretical immunological hy-
pothesis. Our main goal was to present a general
architecture of a TAT based AIS and an immune-
inspired algorithm for anomaly detection that could
deal with temporal events. We presented some pre-
liminary results obtained with artificially generated
data-sets that meet some of the characteristics ob-
served on real-world contextual data-sets. We have
also started to analyse the appropriateness of using
TAT in both a detection and classification context.
Despite the limited diversity of the data-sets used,
we believe that the algorithm proposed show that TAT
possesses a handful of promising properties when ap-
plied to temporal anomaly detection. Firstly, each
environment has its own characteristics and therefore
the detection system should reflect this individuality,
through the automatic adjustment of each cell acti-
vation threshold. Secondly, TCELL activation is an
automatic process based on changes in signal inten-
sity and the current values for the K and P. Each
TCELL has its sensitivity adjusted to a baseline that
is characteristic of the past and current activity. Fi-
nally, in TAT, normal activity is manifested by the
presence of recurrent signals and abnormal activities
correspond to exceptional signals for which the reper-
toire of TCELLs should not be adjusted. This is pre-
cisely what is supposed to happen in the detection of
anomalies in real-world applications.
In this phase we neglected some essential prop-
erties of the natural IS that can make adaptation to
evolving normality and anomaly detection more ro-
bust and reliable: immune memory and clonal dy-
namics. Future developments of the TAT-based de-
tector should aim at incorporating these properties.
Less because this would make the AIS in line with
the natural counterpart, but because variation in clonal
size can be a way of adjusting the weight of each
TCELL specificity in the ”committee”, reflecting not
only the history of the signals but also the history of
co-occurrences of those signals.
The preliminary results obtained are in line with
those described by the authors in (Antunes and Cor-
reia, 2008). The results were also promising and the
ongoing research give us confidence to deploy a TAT-
based algorithm for anomaly detection.
ACKNOWLEDGEMENTS
The authors acknowledge the facilities and research
environment gracefully provided by the CRACS
(Center for Research in Advanced Computing Sys-
tems) research unit, an INESC associate of the Fac-
ulty of Science, University of Porto.
REFERENCES
Antunes, M. and Correia, M. (2008). TAT-NIDS: an
immune-based anomaly detection architecture for net-
work intrusion detection. IWPACBB08 - Advances in
Soft Computing (Springer), pages 6067.
Carneiro, J., Paixo, T., Milutinovic, D., Sousa, J., Leon,
K., Gardner, R., and Faro, J. (2005). Immunological
self-tolerance: Lessons from mathematical modeling.
Journal of Computational and Applied Mathematics,
184(1):77100.
de Castro, L. and Timmis, J. (2002). Artificial Immune Sys-
tems: A New Computational Intelligence Approach.
Springer.
Forrest, S., Perelson, A., Allen, L., and Cherukuri, R.
(1994). Self-nonself discrimination in a computer.
Proceedings of the 1994 IEEE Symposium on Re-
search in Security and Privacy, pages 201212.
Greensmith, J., Twycross, J., and Aickelin, U. (2006). Den-
dritic cells for anomaly detection. In Proc. of the IEEE
World Congress on Computational Intelligence, pages
664671.
Grossman, Z. and Paul, W. (1992). Adaptive cellular in-
teractions in the immune system: The tunable acti-
vation threshold and the significance of subthreshold
responses. Proceedings of the National Academy of
Sciences, 89(21):1036510369.
Kim, J., Bentley, P., Aickelin, U., Greensmith, J., Tedesco,
G., and Twycross, J. (2007). Immune system ap-
proaches to intrusion detection - a review. Natural
Computing, 6(4):413466.
Pedroso, J. (2007). Simple Metaheuristics Using the Sim-
plex Algorithm for Non-linear Programming. LNCS,
4638:217221.
Sompayrac, L. (2008). How the Immune System Works.
Blackwell Publishing.
BIOSIGNALS 2009 - International Conference on Bio-inspired Systems and Signal Processing
362
