
CLOSURE IN ARTIFICIAL CELL SIGNALLING NETWORKS
Investigating the Emergence of Cognition in Collectively
Autocatalytic Reaction Networks
James Decraene
Artificial Life Laboratory, Research Institute for Networks and Communications Engineering
Dublin City University, Glasnevin, Dublin, Ireland
Keywords:
Cell Signalling Networks, Closure, Evolutionary Dynamics, Minimal Cognition.
Abstract:
Cell Signalling Networks (CSNs) are complex biochemical networks responsible for the coordination of cellu-
lar activities in response to internal and external stimuli. We hypothesize that CSNs are subsets of collectively
autocatalytic reaction networks. The signal processing or cognitive abilities of CSNs would originate from
the closure properties of these systems. We investigate how Artificial CSNs, regarded as minimal cognitive
systems, could emerge and evolve under this condition where closure may interact with evolution. To as-
sist this research, we employ a multi-level concurrent Artificial Chemistry based on the Molecular Classifier
Systems and the Holland broadcast language. A critical issue for the evolvability of such undirected and au-
tonomous evolutionary systems is to identify the conditions that would ensure evolutionary stability. In this
paper we present some key features of our system which permitted stable cooperation to occur between the
different molecular species through evolution. Following this, we present an experiment in which we evolved
a simple closed reaction network to accomplish a pre-specified task. In this experiment we show that the
signal-processing ability (signal amplification) directly resulted from the evolved systems closure properties.
1 INTRODUCTION
Cell Signalling Networks (CSNs) are biochemical
networks of interacting molecules (proteins, ions, sec-
ondary messengers, etc) occurring in living cells.
Through complex molecular interactions (e.g., sig-
nal transduction), CSNs are able to coordinate critical
cellular activities (e.g., cell differentiation, apoptosis)
in response to internal and external stimuli.
In the presence of a genetic subsystem, the pro-
duction of CSN components may result from the
translation of the genetic code. However to coordi-
nate this decoding process, a newborn cell still re-
quires a minimal CSN which would be inherited from
the parent cell during cellular division. In a prebiotic
world, in which genetic material may potentially not
be present, “primitive” CSNs would need an alterna-
tive mechanism to ensure their production and main-
tenance.
We may argue that both of these mini-
mal/primitive CSNs from the post/prebiotic world
may have to replicate themselves prior to the cellular
division. This would allow the replicated CSNs to be
“distributed” to the offspring cells. Errors may occur
during the replication process, e.g., an offspring
cell may inherit only a partial or mutant CSN. Thus
resulting in potentially defective cells which would
lead to a variety of undesired effects (e.g., premature
cell death).
As a result, the “fitness” of a cell is implicitly rep-
resented by the survival and performance of a cell in
achieving self-maintenance and cell-level replication.
Based on the above assumption, we hypothesize that
CSNs may be regarded as subsets of collectively au-
tocatalytic (closed) reaction networks. In these closed
and synergetic reaction networks, self-maintenance is
achieved through the systems ability to produce the
catalysts and substrates for all its reactions. This cat-
alytic closure property originates from the autocat-
alytic set theory, which is one of the current candi-
date theories explaining the origins of life (Kauffman,
1997).
These closed reaction networks would have the
additional ability to replicate themselves as a whole
(during cellular division). Through the evolutionary
process, we suggest that these systems may develop
signal processing abilities which are typical of real
CSNs. However these Artificial CSNs (ACSNs) must
preserve closure to self-maintain and survive. Thus
the systems signal-processing abilities may originate
107
Decraene J. (2009).
CLOSURE IN ARTIFICIAL CELL SIGNALLING NETWORKS - Investigating the Emergence of Cognition in Collectively Autocatalytic Reaction Networks.
In Proceedings of the International Conference on Bio-inspired Systems and Signal Processing, pages 107-114
DOI: 10.5220/0001545301070114
Copyright
c
SciTePress

from the closure properties where inputs (internal or
external stimuli) are perturbations to the system. The
system outputs would result in a modification of the
systems state and of its closure properties.
Following (van Duijn et al., 2006) we regard the
coupling of these ACSNs signal-processing abilities
and closure properties as minimal principles for life
and cognition. By examining the conditions for the
emergence of distinctive ACSNs signal-processing
abilities, we investigate the emergence and origins of
minimal cognition. These ACSNs may thus be re-
garded as Artificial CognitiveSystems which may po-
tentially provide a range of applications in computing
and engineering. Thus the goals of this research are
twofold and can be summarized as follows:
• From a theoretical point of view: to study the ori-
gins and complexity evolution of cognition in bi-
ological systems.
• From a practical perspective: to provide a new
computational paradigm inspired by nature.
To assist this research we developed the MCS.bl,
a string-based Artificial Chemistry (AC) based on
the Molecular Classifier Systems (MCS) (McMullin
et al., ) and the Holland broadcast language (Hol-
land, 1992). This technique examines the evolution
of biochemical networks from an undirected, self-
engineered and autonomous perspective. In this ap-
proach no explicit fitness function is defined, the evo-
lutionary process is driven by the capacity of CSNs
to self-maintain and perform the necessary signal-
processing operations or “computation” to achieve
cell-level replication.
A key step of this research is to identity the
conditions which provide evolutionary stability to
the closed reaction networks when simulated in the
MCS.bl. Examining such phenomena relates closely
to other studies which have been conducted on Hol-
land’s Alpha-Universes (Holland, 1976), Tierra (Ray,
1991) and Alchemy (Fontana and Buss, 1994b). In
this paper we first describe the MCS.bl. We then
discuss how a multi-level and concurrent model en-
hanced the evolutionary stability of the system. Fi-
nally we present an experiment in which we evolved
a minimal closed ACNS to accomplish a pre-specified
task (signal amplification).
2 MOLECULAR CLASSIFIER
SYSTEMS
Molecular Classifier Systems are a class of string-
rewriting based AC inspired by Learning Classifier
Systems (LCS). As opposed to traditional string-
rewriting systems, operations are stochastic and re-
flexive (no distinction made between operands and
operators). The behaviour of the condition (binding)
properties and action events (enzymatic functions) is
defined by a language specified within the MCS. This
“chemical” language defines and constrains the com-
plexity of the chemical reactions that may be mod-
elled and simulated. In this AC, all reactants are cat-
alytic in the sense that they are not consumed dur-
ing reactions. These reactions result from successful
molecular interactions which occur at random. When
a reaction occurs, a product molecule is inserted into
the reactor.
We proposed a simplification of the Holland
broadcast language (Decraene et al., 2007) which is
used as the MCS chemical language resulting in the
MCS.bl system. The MCS.bl differs from the original
MCS (McMullin et al., ) by introducing more com-
plex chemical reactions (only replications may occur
in the MCS). A molecule may contain several condi-
tion/action rules which define the binding and enzy-
matic properties. A reaction between molecules oc-
curs if at least one conditional part from any rules in
a molecule A matches a target molecule B. A is re-
garded as an enzyme whereas B is regarded as a sub-
strate molecule. When a reaction occurs, the action
part from the satisfied rule in A is utilized to perform
the enzymatic operations upon the bound substrate
molecule B. This operation results in the production
of another offspring (product). If several rules in A
are satisfied by B, then one of these rules is picked
at random and employed to carry out the enzymatic
function.
A number of differences exist between our sim-
plified broadcast language (BL) and the LCS, e.g.,
the LCS’s alphabet is λ = { 1, 0, #} whereas the
BL includes additional symbols Λ = {1, 0, ∗, :
, ♦, △,
′
, ▽}. The basic elements of the BL are
strings made from Λ called broadcast devices. A
broadcast device is parsed into zero, one or more
broadcast units, where each unit represents a single
condition/action rule. The symbol ∗ separates broad-
cast units within a broadcast device. The symbol :
separates a condition from an action within a sin-
gle broadcast unit. 0s and 1s are basic informational
symbols. {♦, ▽, △} are single/multiple character(s)
wildcards that may also transpose matched strings
into output strings. Quoted symbols (preceded by
′
)
are prevented from interpretation. Figure 1 depicts an
example broadcast device which may bind and react
with a copy of itself, this reaction is presented in Fig-
ure 2 .
A detailed description is omitted in this paper, see
BIOSIGNALS 2009 - International Conference on Bio-inspired Systems and Signal Processing
108

Figure 1: An example broadcast device.
Figure 2: Example reaction.
Table 1: Example operations realized with the MCS.bl.
Enzyme substrate product operation
∗▽1 : ▽0 1 : 0
/
0
no reaction
∗▽1 :
′
∗ ▽ 0 : 1 ∗0 : 1 activation
∗
′
∗ 0▽ : 0▽ ∗0 : 1 0 : 1 inhibition
∗▽ : ▽ ∗00 : 11 ∗00 : 11 universal replication
∗▽0 : ▽0 ∗▽0 : ▽0 ∗▽0 : ▽0 self-replication
∗▽1 : ▽10 ∗0 : 1 ∗0 : 10 concatenation
∗▽1 : ▽ ∗0 : 1 ∗0 : cleavage
(J.Decraene, 2006) for full specification of our BL im-
plementation. Table 1 presents a number of example
operations that can be realized with the MCS.bl.
3 EXAMINING EVOLUTIONARY
STABILITY
As this evolutionary system is an undirected ap-
proach, the first key step was to obtain catalytic net-
works that are able to self-sustain over time. In order
to achieve such evolutionary robustness, it is neces-
sary for these networks to possess mechanisms which
provide some protection from parasites that may be
formed. In this section we examine the system evolu-
tionary stability firstly when implemented as a single
level selectional model, secondly when the MCS.bl is
implemented as a multi-level concurrent system.
3.1 Single-level Selectional Model
In a single-level selectional model, all molecules are
contained in a single reactor in which they are com-
peting with each other. This molecular competition is
referred to as the first level of selection. Each time a
successful reaction occurs, a product molecule is in-
serted into the reactor, if the latter is saturated then an-
other molecule (selected at random) is removed from
the reactor space (designating the system outflow),
see Figure 3.1.
A formal explanation of this outcome is given by
modelling the system with the (approximate, continu-
ous) catalytic network equation (Stadler et al., 1993).
The state of the system is described by the concentra-
tion vector x = (x
1
, . . . , x
n
) with x
1
+ . . . + x
n
= 1 and
x
i
> 0, where x
i
refers to the concentrationof a molec-
ular species (or collection of “chemically equivalent”
species) s
i
. The general dynamic behaviour is then
given by:
˙x
k
=
n
∑
i=1
n
∑
j=1
α
k
ij
x
i
x
j
− x
k
n
∑
i, j,l=1
α
l
ij
x
i
x
j
(1)
with k = 1, . . . , n
α
k
ij
are the rate constants for each reaction s
i
+
s
j
→ s
i
+ s
j
+ s
k
. In here, these simplify to:
α
k
ij
=
1 if s
i
+ s
j
→ s
i
+ s
j
+ s
k
0 otherwise
(2)
Previous studies (Decraene et al., 2008) demon-
strated that the spontaneous emergence and self-
maintenance of replicases in the MCS.bl was unlikely
to occur. Consequently experiments were conducted
in which, as in the Tierra system, a hand-designed
molecule called the ancestor was introduced into
the population. This replicase molecule was hand-
designed so as to possess a high binding specificity,
thus preventing exploitation from other molecules.
This specificity ensures a rapid dominance over a ran-
domly generated molecular population (which would
not be possible if a universal replicase, having no
binding specificity, was employed instead; see (De-
craene et al., 2008)). Various molecular mutations
through successful reactions were possible and a con-
straint over the molecular complexity was set due to
computational limitations (experimental details and
mathematical analysis may be found in (Decraene
et al., 2008)). Although this MCS.bl implementation
CLOSURE IN ARTIFICIAL CELL SIGNALLING NETWORKS - Investigating the Emergence of Cognition in
Collectively Autocatalytic Reaction Networks
109
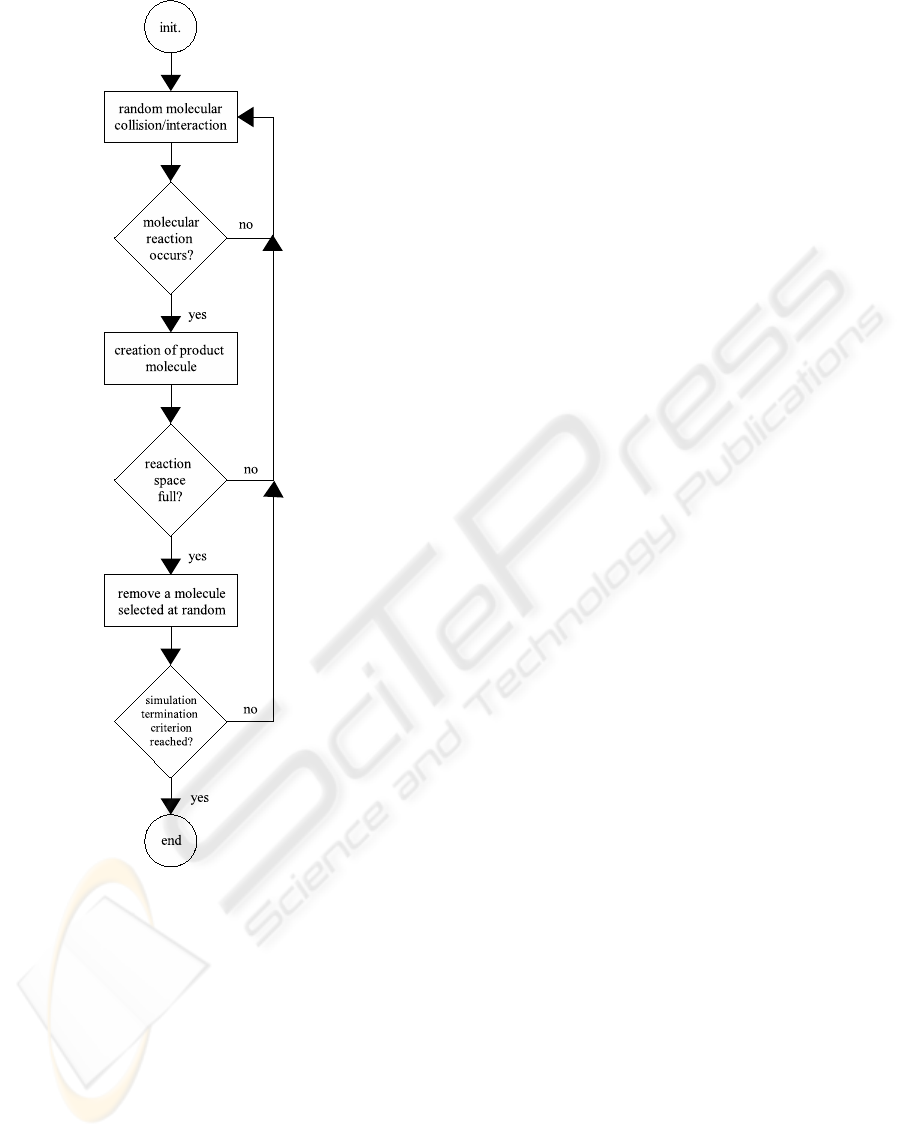
Figure 3: Flowchart of the single-level selectional model.
was to some extent similar to the Tierra system, we
did not observe the emergence of collectively auto-
catalytic networks. Instead we observed a degenerate
outcome characterized by an “elongation catastrophe”
phenomenon:
During evolution, a series of parasitic displace-
ments was observed leading to ever longer molecules.
However, as the mutation effect increases (with the
length of a molecule), disruptive structural changes
may also occur more frequently. We observed
that consequences of these structural changes were
twofold:
• Molecules may become inactive, thus being con-
sidered as substrate or waste molecules. These
waste molecules would then accumulate and pre-
vent any further enzymatic activities. A potential
consequence of the BL syntax is that a mutation
may disrupt dramatically the behaviour of an en-
zymatic molecule. This degenerative effect may
be regarded as the consequence of the BL “brittle-
ness” (asymmetric mutation effects (Ray, 1992)).
• The binding specificity may be increased. Mu-
tations leading to the insertion of informational
symbols such as 0s and 1s may also increase sig-
nificantly the binding specificity of molecules. As
a result, although these molecules may still pos-
sess an active site capable of some enzymatic
function, they could not catalyse any molecules
in the reactor.
Both of these phenomena result in a continual de-
crease in the overall reaction rate until reactions cease
completely (i.e., the system decays and ultimately
reaches a state of stasis). These results obtained with
this single-level AC exhibited unexpected evolution-
ary dynamics which resulted in various degenerate
cases: No stable cooperation between the molecular
species could be observed in the evolutionary simula-
tions. Thus this first implementation of our evolution-
ary system suffered from a critical lack of “robust-
ness”.
3.2 Multi-level Selectional and
Concurrent Model
In the multi-level selectional model, we introduce
multiple reactors where each of them contains a pop-
ulation of molecules. These reactors or cells may be
subjected to cellular division, which results in the re-
placement of the parent cell and creation of two off-
spring cells. However, the number of cells in the uni-
verse is fixed. As a result such a cellular division also
triggers the removal of another cell selected at ran-
dom. In a similar manner to molecules, cells are com-
peting with each other which is regarded as the second
level of selection.
In contrast to the single level model, successful
reactions do not lead to the removal of a random
molecule in the reaction space. Thus the number m
of molecules contained in a cell may increase until
the cell is full (i.e., when m is equal to the cell max-
imum capacity c). When a cell is full, a division oc-
curs as follows: Half of the molecules contained in
the cell are selected at random, then these molecules
are removed from this cell and are inserted into the
offspring cell. This newly created cell is then inserted
into the cellular population. Finally, a cell is picked at
BIOSIGNALS 2009 - International Conference on Bio-inspired Systems and Signal Processing
110
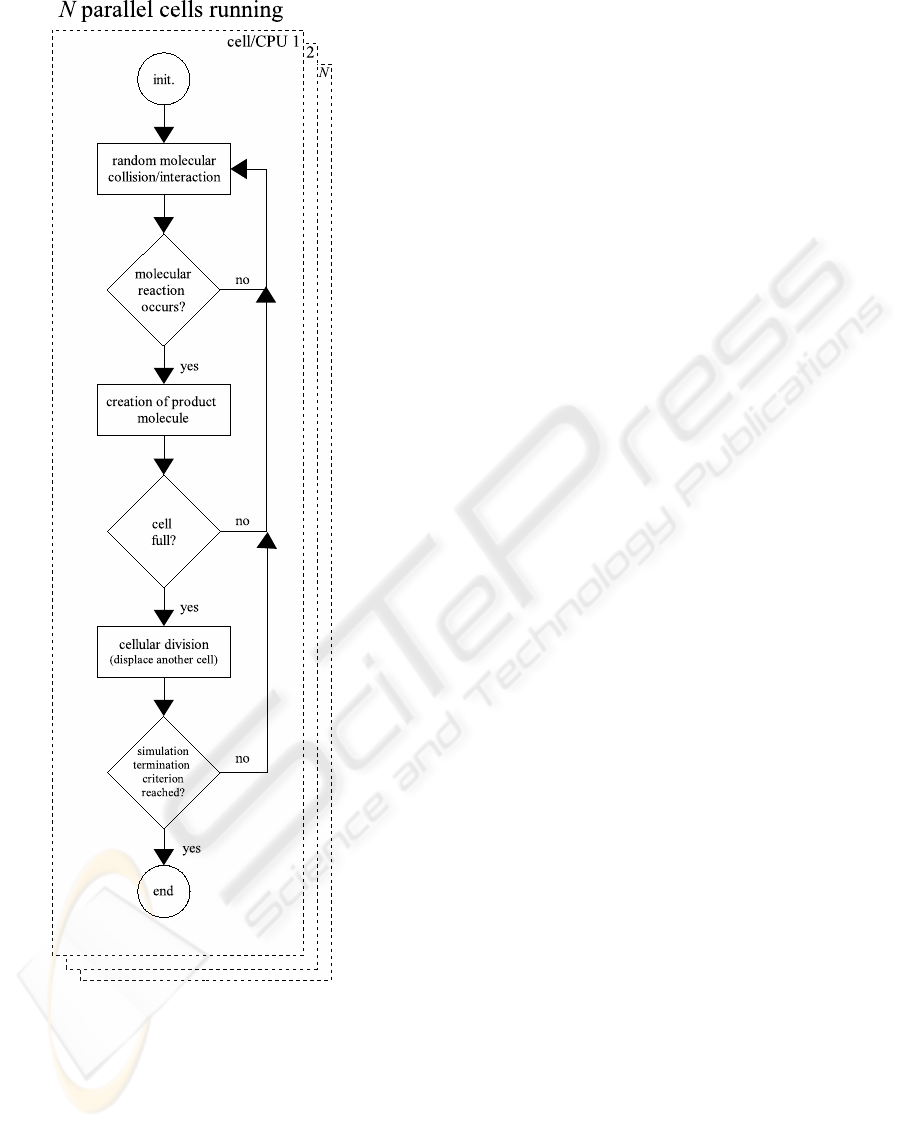
Figure 4: Flowchart of the multi-level and concurrent
model.
random (other than the offspring and parent cell) and
removed from the cell population, see Figure 3.2.
Furthermore this multi-level model was imple-
mented as a concurrent system where each cell is run
on a single CPU. In this concurrent model, the fittest
cells would not only be the cells that exhibit a high
rate of successful reactions (when compared to the to-
tal number of molecular collisions), but also cells that
contain molecules that are fast to compute. For exam-
ple let us consider two cells containing complete re-
action networks (i.e., all molecular collisions lead to
the successful production of molecules). Those cells
would moreover contain molecules having different
computational complexities. In here the cell which
possesses a smaller overall molecular computational
complexity will have the selective advantage. This
computational complexity introduces in our model a
notion of chemical kinetics and may alter the cellular
growth rate (i.e., the cells fitness).s
We extended the MCS.bl with the multi-level con-
current model described above, our results indicated
that none of the evolved cells resulting from the sim-
ulations suffered from elongation catastrophe. The
nature of the evolved populations resulting from the
simulation runs were equivalent to those expected
from systems such as Alchemy: We observed the
rapid domination of molecular organizations which
involved a limited number of replicases capable to
self-sustain over time.
Moreover, in traditional multi-level selectional
models infected cells would usually be displaced only
when these have decayed (Cronhjort and Blomberg,
1997; Hogeweg and Takeuchi, 2003). Whereas in
our multi-level concurrent AC, infected cells (having
a smaller growth rate) would rapidly be displaced by
healthier cells (having a higher growth rate) due to
the concurrentnature of the system. This concurrency
property increased the system’s ability to control par-
asitism.
4 EVOLVING CLOSED
REACTION NETWORKS
The previous examination on evolutionary stabil-
ity provided us with the necessary understanding to
evolve closed reaction networks that are able to self-
sustain over time. These closed reaction networks
are regarded as primitive ACNS. Preliminary experi-
ments suggested that when self-replication is enabled,
evolved networks would present only limited interest.
As these networks would converge toward molecu-
lar organizations involving a small number of altru-
ist replicases. In the following experiment we dis-
able self-replication, and employ a minimalist (hand-
designed) collectively autocatalytic reaction network,
see Figure 5.A. Let S1 = ∗▽0 : ▽1, S2 = ∗▽0 : ▽0,
S3 = ∗▽1 : ▽1, S4 = ∗▽1 : ▽1, S5 = ∗▽⋄ : ▽1 and
S6 = ∗▽⋄ : ▽0.
In section 3.2, a cell would divide only when sat-
CLOSURE IN ARTIFICIAL CELL SIGNALLING NETWORKS - Investigating the Emergence of Cognition in
Collectively Autocatalytic Reaction Networks
111
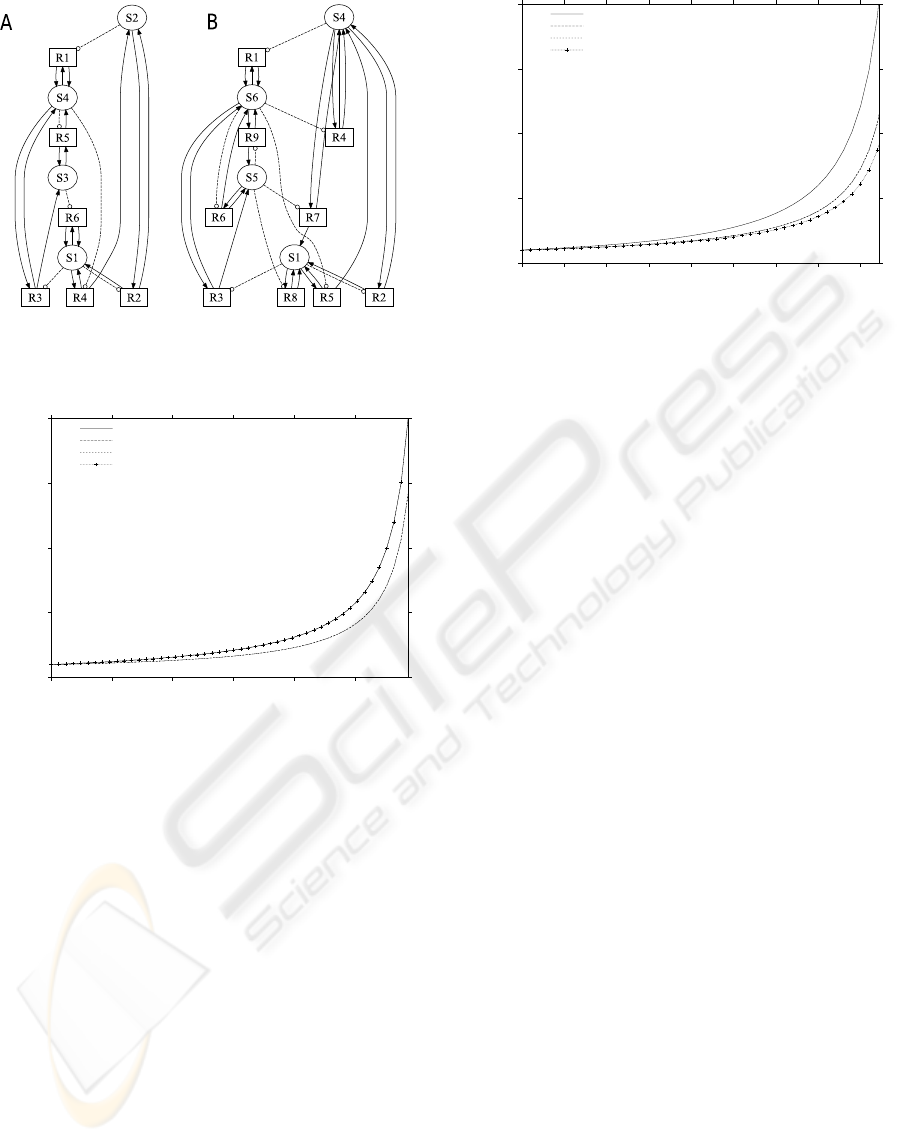
Figure 5: A: Seed closed reaction network. B: Evolved
closed reaction network promoting growth of molecular
species S1.
0
50
100
150
200
0 0.01 0.02 0.03 0.04 0.05
Number of molecules
Time
S1
S2
S3
S4
Figure 6: Dynamics of seed closed reaction network, the
species S1 and S4 overlapping (top line) and S2 and S3 also
overlapping.
urated. We define a new cellular division criterion:
Here a cell would divide when the amount of molec-
ular species S1 reaches 200. The cellular growth rate
now depends on the molecular growth rate of S1. The
ability of the closed reaction network to promote the
growth of S1 defines the cells fitness (i.e., a cell would
increase its fitness by exhibiting a higher S1 growth
rate ). Thus the pre-specified task assigned to these
simple closed ACSNs is to amplify the “signal” S1.
We may also regard S1 as a necessary molecule such
as a membrane molecule to allow the cellular division
to occur. We first present the dynamics of the seed
closed reaction network when the initial amount of
each molecular species is equal to 10, see Figure 6.
This graph was obtained by solving the ODE system
generated by the reaction network (in SBML format)
using the SBML ODE solver (Machn´e et al., 2006).
An evolutionary simulation was conducted in
which 31 concurrent cells were employed. These
0
50
100
150
200
0 0.005 0.01 0.015 0.02 0.025 0.03 0.035 0.04
Number of molecules
Time
S1
S5
S6
S4
Figure 7: Dynamics of evolved closed reaction network,
the species S5 and S6 overlapping (middle line), S1 is the
top line and S4 is the bottom line. Each molecular species
amount is initialized to 10.
cells were seeded with the molecular species (with
initial amount of molecules 1) depicted in Figure 5.A.
This experiment was run for one hour, on average
the number of interactions per cell per hour was over
4.10
6
. The resulting dominant molecular organiza-
tion is depicted in Figure 5.B. We first observed that
this evolved network successfully maintained closure.
We note that both species S2 and S3 have been re-
placed by new molecular species S5 and S6. These
new molecular species increased the overall number
of reactions from 6 to 9. We compare the dynamics of
this evolved closed reaction network against the seed
closed reaction network, see Figure 7.
In Figure 7, it can be seen that S1 reaches the di-
vision threshold at t ≈ 0.0422 whereas in the seed
closed reaction network S1 would reach this threshold
at t ≈ 0.0588. By reaching this threshold at an earlier
stage, the evolved network gained a selective advan-
tage over the seed network. We observed the emer-
gence of molecular species S5 and S6 which had for
effect to promote the growth of S1 whilst maintaining
closure. The network closure properties evolved and
allowed the network to promote the growth of species
S1.
Moreover results indicated that when a cellular
division occurs, the amount of the different molecu-
lar species would be optimized, see Figure 8. This
optimization resulted in the ability to encourage the
growth of S1 while limiting the growth of species S4,
S5 and S6. However this amount distribution would
not lead to the displacement of S4, S5 and S6 as these
species are necessary to maintain closure.
In Figure 8, it can be observed that S1 would reach
the division threshold when t ≈ 0.00973. This op-
timized amount distribution provided this network a
selective advantage over networks having differing
BIOSIGNALS 2009 - International Conference on Bio-inspired Systems and Signal Processing
112
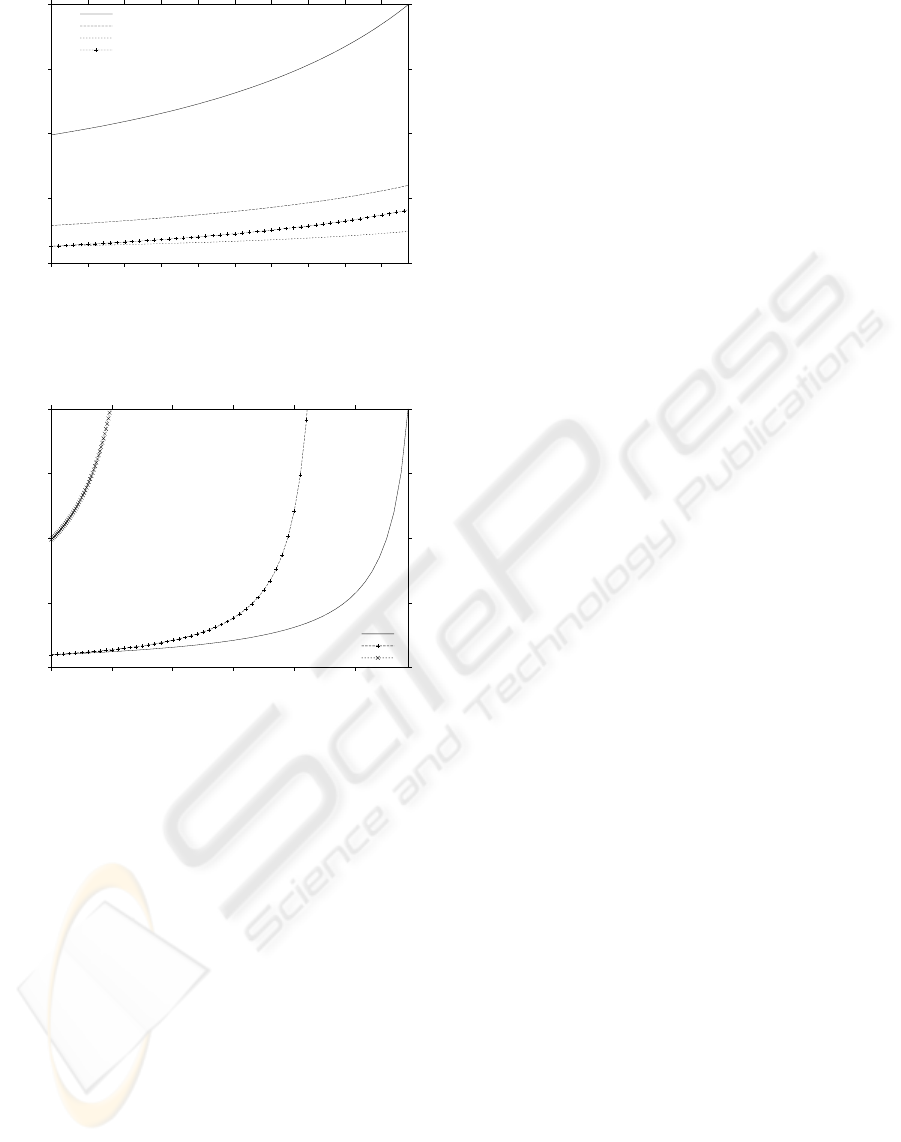
0
50
100
150
200
0 0.001 0.002 0.003 0.004 0.005 0.006 0.007 0.008 0.009
Number of molecules
Time
S1
S5
S6
S4
Figure 8: Typical dynamics of an evolved closed reaction
network with optimized initial molecular amounts resulting
from a cellular division
0
50
100
150
200
0 0.01 0.02 0.03 0.04 0.05
Number of molecules
Time
S1
a
S1
b
S1
c
Figure 9: Comparison of S1 dynamics with differing net-
works and initial molecular amount distributions. S1
a
and
S1
b
depicts the dynamics of S1 with initial molecular 10, us-
ing the seed reaction network and evolved reaction respec-
tively. S1
c
represents the dynamics of S1 using the evolved
reaction network and optimal initial molecular amount dis-
tribution.
amount distributions (as shown in Figure 7).
Finally Figure 9 compares the different dynamics
of S1 using the different networks and initial number
of molecules. This comparison highlights the succes-
sive improvements in the networks fitness (i.e., has-
tened the production of molecules A).
5 DISCUSSION
We now discuss the current limitations of our pro-
posed evolutionary system and how future work may
address these issues. In section 4, we presented a
closed reaction network which was evolved to carry
out a pre-specified task. We examined a second
set of identical evolutionary simulations to examine
whether alternativeevolutionary trajectories could oc-
cur. In all of these experiments, we observed the con-
vergence(from a phenotypic point of view) of the sys-
tem towards reaction networks similar to Fig. 5.B.
Moreover the evolutionary process would only in-
volve a limited number of incremental steps in fitness.
Based on these observations we may express that our
proposed AC still lacks some evolvability and failed
to display a “remarkable” evolution of complexity.
Some may argue that the current system is sim-
ply not complex enough and should incorporate more
realistic features such as mass conservation, multi-
dimensional space, a genetic code or advanced chem-
ical kinetics. These complementary properties would
certainly broaden the complexity of an already dif-
ficult and incomplete investigation. However there
would be no guarantee of improved results, i.e., ex-
hibiting a more interesting evolutionary growth of
complexity: A first reason for this assertion is that the
impact of environmental constraints on the evolution
of complexity still remains to date an open question
(Gershenson and Lenaerts, 2008). Secondly there is
currently no theoretical framework that would lead to
desired evolutionary growth of complexity with con-
fidence.
We hypothesise that as long there is no such the-
oretical frameworks for the study of ACs, the lat-
ter would remain a black art approach where unex-
pected results are observed. Therefore a minimal-
ist/simplified approach to ACs, where the system is
still analytically tractable and examined using current
mathematical methods, should be adopted. In keeping
with this, our proposed AC contributed, to some ex-
tent, to the understanding of minimalist ACs and their
evolutionary dynamics.
Future work would benefit from further theoreti-
cal research, it could involve the development of For-
mal Methods for Artificial Life (AL), this would fa-
cilitate the comparison of differing ACs. Such a for-
mal approach to AL had already been initiated but
was discontinuated (Grogono et al., 2003), an alterna-
tive could be to extend the AC formalism. Moreover
the development of further analytical tools such as the
Organization Theory, which was initiated by (Fontana
and Buss, 1994a) and later enriched by (Dittrich and
Speroni, 2007), would as well assist and contribute
to the understanding and ultimately engineering of
Complex Adaptive Systems using ACs.
Finally, although our approach may present some
limitations in terms of evolutionary growth of com-
plexity, it still demonstrated the successful evolu-
tion of closed reaction networks to accomplish a pre-
specified task. This may be regarded as an initial
CLOSURE IN ARTIFICIAL CELL SIGNALLING NETWORKS - Investigating the Emergence of Cognition in
Collectively Autocatalytic Reaction Networks
113

promising result demonstrating closure in CSNs and
may suggest a novel method to engineer autonomous
Complex Adaptive Systems for real-world applica-
tions.
6 CONCLUSIONS
We discussed how CSNs could be considered as sub-
sets of closed reaction networks and the potential role
of closure on CSNs evolution. We presented our evo-
lutionary system: a multi-level concurrent Artificial
Chemistry based on the Molecular Classifier Systems
and the Holland broadcast language. The develop-
ment of this multi-level and concurrent model was
motivated to improve evolutionary stability, which
was a key missing feature of the single-level MCS.bl.
We then presented these different properties which
permitted the systems robustness to be ameliorated.
We finally presented an experiment in which a sim-
ple closed reaction network was successfully evolved
and optimized to carry out a pre-specified task (signal
amplification) whilst maintaining closure. This opti-
mized signal-processing ability directly resulted from
the evolved closure properties. We finally discussed
the contributions and future directions of this work.
ACKNOWLEDGEMENTS
We acknowledge the ESIGNET (Evolving Cell Sig-
nalling Networks in Silico) Project funding (contract
no. 12789).
REFERENCES
Cronhjort, M. and Blomberg, C. (1997). Cluster compart-
mentalization may provide resistance to parasites for
catalytic networks. Physica D: Nonlinear Phenom-
ena, 101(3-4):289–298.
Decraene, J., Mitchell, G. G., and McMullin, B. (2007).
Evolving artificial cell signaling networks: Perspec-
tives and methods. In Advances in Biologically In-
spired Information Systems, volume 69, pages 165–
184. Springer.
Decraene, J., Mitchell, G. G., and McMullin, B. (2008).
Unexpected Evolutionary Dynamics in a String-Based
Artificial Chemistry. Proceedings of Artificial Life XI,
MIT Press.
Dittrich, P. and Speroni, P. (2007). Chemical Organi-
sation Theory. Bulletin of Mathematical Biology,
69(4):1199–1231.
Fontana, W. and Buss, L. (1994a). The arrival of the fittest:
Toward a theory of biological organization. Bulletin
of Mathematical Biology, 56(1):1–64.
Fontana, W. and Buss, L. (1994b). What Would be Con-
served if “the Tape were Played Twice”? Proc. of the
National Academy of Sciences, 91(2):757–761.
Gershenson, C. and Lenaerts, T. (2008). Evolution of Com-
plexity. Artificial Life, 14(3):241–243.
Grogono, P., Chen, G., Song, J., Yang, T., and Zhao, L.
(2003). Laws and life. In Proceedings of the 7th
IASTED Conference on Artificial Intelligence and Soft
Computing (ASC 2003), pages 158–163.
Hogeweg, P. and Takeuchi, N. (2003). Multilevel selection
in models of prebiotic evolution: compartments and
spatial self-organization. Origins of Life and Evolu-
tion of the Biosphere, 33(4-5):375–403.
Holland, J. (1976). Studies of the spontaneous emergence
of self-replicating systems using cellular automata and
formal grammars. Automata, Languages, Develop-
ment, pages 385–404.
Holland, J. (1992). Adaptation in natural and artificial sys-
tems. MIT Press Cambridge, MA, USA.
J.Decraene (2006). The Holland Broadcast Language.
Technical Report ALL-06-01, RINCE, Dublin City
University. http://elm.eeng.dcu.ie/˜alife/jd/ALL-06-
01/.
Kauffman, S. (1997). At Home in the Universe. Mathemat-
ical Social Sciences, 33(1):94–95.
Machn´e, R., Finney, A., M¨uller, S., Lu, J., Widder, S., and
Flamm, C. (2006). The sbml ode solver library: a na-
tive api for symbolic and fast numerical analysis of re-
action networks. Bioinformatics, 22(11):1406–1407.
McMullin, B., Kelly, C., and O’Brien, D. Multi-level se-
lectional stalemate in a simple artificial chemistry. In
Proceedings of ECAL 2007, LNCS. Springer.
Ray, T. (1991). An approach to the synthesis of life. Artifi-
cial Life II, 10:371–408.
Ray, T. (1992). Evolution, ecology and optimization of dig-
ital organisms. Santa Fe.
Stadler, P. F., Fontana, W., and Miller, J. H. (1993). Random
catalytic reaction networks. Phys. D, 63(3-4):378–
392.
van Duijn, M., Keijzer, F., and Franken, D. (2006).
Principles of Minimal Cognition: Casting Cognition
as Sensorimotor Coordination. Adaptive Behavior,
14(2):157.
BIOSIGNALS 2009 - International Conference on Bio-inspired Systems and Signal Processing
114
