
Nash Equilibria in Multi-Agent Swarms
Carsten Hahn, Thomy Phan, Sebastian Feld, Christoph Roch, Fabian Ritz, Andreas Sedlmeier,
Thomas Gabor and Claudia Linnhoff-Popien
Mobile and Distributed Systems Group, LMU Munich, Munich, Germany
carsten.hahn, thomy.phan, sebastian.feld, christoph.roch, fabian.ritz, andreas.sedlmeier, thomas.gabor,
Keywords:
Multi-Agent Systems, Reinforcement Learning, Nash Equilibrium, Partial Observability, Scaling.
Abstract:
In various settings in nature or robotics, swarms offer various benefits as a structure that can be joined easily
and locally but still offers more resilience or efficiency at performing certain tasks. When these benefits are
rewarded accordingly, even purely self-interested Multi-Agent reinforcement learning systems will thus learn
to form swarms for each individual’s benefit. In this work we show, however, that under certain conditions
swarms also pose Nash equilibria when interpreting the agents’ given task as multi-player game. We show
that these conditions can be achieved by altering the area size (while allowing individual action choices) in
a setting known from literature. We conclude that aside from offering valuable benefits to rational agents,
swarms may also form due to pressuring deviants from swarming behavior into joining the swarm as is typical
for Nash equilibria in social dilemmas.
1 INTRODUCTION
Flocking behavior can be observed in many species
in nature. For example fish or birds coordinate their
actions in order to form a swarm. This yields ben-
efits like: hydrodynamic efficiency, higher mating
chances, enhanced foraging success, enhanced preda-
tor detection, decreased probability of being caught
and overall reduced individual effort. In order to profit
from such benefits, swarming has also been trans-
ferred to technical systems featuring multiple robots
or drones (Brambilla et al., 2013; Christensen et al.,
2015). Swarms can be especially useful when they
allow to use smaller, simpler and effectively cheaper
robots and use the swarm to still allow for complex
behavior and error resilience (against communication
failures, bugs or external influences). However, since
swarms rely on the emergent behavior of a group of
individual agents, swarm behavior is often hard to
pre-program or even just predict (Pinciroli and Bel-
trame, 2016).
In this paper, we consider the issue of swarms con-
sisting of self-adaptive, learning agents and raise the
question under which conditions these agents tend to
form swarms purely out of self-interest. Here, mul-
tiple agents coming together within a swarm in the
first place is not given or programmed but on its own
already an emergent behavior.
¨
Ozg
¨
uler and Yıldız
(
¨
Ozg
¨
uler and Yıldız, 2013) introduced a theoretical
model to examine how swarms form. They mod-
eled foraging swarm behavior as a non-cooperative N-
player game and have shown that the resulting swarms
pose a Nash equilibrium.
Hahn et al. (Hahn et al., 2019) have considered a
continuous predator-prey scenario where a swarm of
agents (resembling fishes) aims to survive for as long
as possible in the presence of an enemy agent (resem-
bling a shark, e.g.). Under certain conditions, swarms
in such Multi-Agent system can emerge solely by
training (using reinforcement learning, e.g.) each
agent on the purely self-interested goal of securing
its own survival. It has been observed that the agents
learn to form clusters because the predator can be dis-
tracted by multiple agents in its vicinity, which in-
creases the survival chance of any individual. In their
work only the prey agents are actively trained while
the present predator follows a predefined static heuris-
tic strategy. The prey agents are self-interested and
maximize solely their own reward (i.e., surviving as
long as possible). The group of agents is trained by
iteratively training only one of the prey agents and
copying its learned policy to all other homogeneous
agents.
The work of (Hahn et al., 2019) focuses mainly
on the examination of the resulting swarms and their
comparison to existing related swarm approaches
234
Hahn, C., Phan, T., Feld, S., Roch, C., Ritz, F., Sedlmeier, A., Gabor, T. and Linnhoff-Popien, C.
Nash Equilibria in Multi-Agent Swarms.
DOI: 10.5220/0008990802340241
In Proceedings of the 12th International Conference on Agents and Artificial Intelligence (ICAART 2020) - Volume 1, pages 234-241
ISBN: 978-989-758-395-7; ISSN: 2184-433X
Copyright
c
2022 by SCITEPRESS – Science and Technology Publications, Lda. All rights reserved

(Reynolds, 1987; Morihiro et al., 2008). But they also
investigate origin and characteristics of the social be-
havior between prey agents and hint that the swarm
behavior of multiple self-interested agents trained us-
ing Multi-Agent reinforcement learning is linked to
Nash Equilibria.
We build on this foundation to further investigate
Nash equilibria in Multi-Agent swarms resulting from
Multi-Agent reinforcement learning. We adopt the
scenario of (Hahn et al., 2019) for our research and
further examine the conditions under which forming
a swarm pays off for each individual agent or un-
der which running individually is the superior strat-
egy. We show that the partial observable scenario
can be expanded and the learned policies can adapt
without any re-training. We relax the imposed crite-
ria on agent homogeneity, i.e., we allow each agent to
choose for itself if it wants to join a swarm or roam on
its own. We observe that this decision poses a social
dilemma as swarms only offer a benefit at a certain
size. However, we also show that some swarm con-
figurations form Nash equilibria under certain condi-
tions, which means that even when swarming might
not be the strictly superior strategy in a certain situa-
tion, deviating from an already instituted is still worse
for a single agent.
2 FOUNDATIONS
2.1 Reinforcement Learning
Reinforcement Learning (RL) is a machine learning
paradigm which models an autonomous agent that
has to find a decision strategy in order to solve a
task. The problem is typically formulated as Markov
Decision Process (MDP) (Howard, 1961; Puterman,
2014) which is defined by a tuple M = hS, A, P , R i,
where S is a set of states, A is the set of actions,
P (s
t+1
|s
t
, a
t
) is the transition probability function and
R (s
t
, a
t
) is the scalar reward function. We assume
that s
t
, s
t+1
∈ S , a
t
∈ A, r
t
= R (s
t
, a
t
), where s
t+1
is
reached after executing a
t
in s
t
at time step t. Π is the
policy space.
The goal is to find a policy π : S → A with π ∈ Π,
which maximizes the expected (discounted) return G
t
at state s
t
for a horizon h:
G
t
=
h−1
∑
k=0
γ
k
· R (s
t+k
, a
t+k
) (1)
where γ ∈ [0, 1] is the discount factor.
A policy π can be evaluated with a value function
Q
π
(s
t
, a
t
) = E
π
[G
t
|s
t
, a
t
], which is defined by the ex-
pected return when executing a
t
at state s
t
and fol-
lowing π afterwards (Bellman, 1957; Howard, 1961).
π is optimal if Q
π
(s
t
, a
t
) ≥ Q
π
0
(s
t
, a
t
) for all s
t
∈ S ,
a
t
∈ A, and all policies π
0
∈ Π. The optimal value
function, which is the value function for any optimal
policy π
∗
, is denoted as Q
∗
and defined by (Bellman,
1957):
Q
∗
(s
t
, a
t
) = r
t
+γ
∑
s
0
∈S
P(s
0
|s
t
, a
t
)·max
a
0
∈A
{Q
∗
(s
0
, a
0
)}
(2)
When Q
∗
is known, then π
∗
is defined by π
∗
(s
t
) =
argmax
a
t
∈A
{Q
∗
(s
t
, a
t
)}.
Q-Learning is a popular RL algorithm to approx-
imate Q
∗
from experience samples (Watkins, 1989).
In the past few years, Q-Learning variants based on
deep learning, called Deep Q-Networks (DQN), have
been applied to high dimensional domains like video
games and Multi-Agent systems (Mnih et al., 2015;
Hausknecht and Stone, 2015; Leibo et al., 2017).
2.2 Game Theory and Multi-Agent
Reinforcement Learning
Multi-Agent Reinforcement Learning (MARL) prob-
lems can be formulated as stochastic game M =
hS , A, P , R , Z, O, ni, where hS ,A, are P equivalently
defined as in MDPs. n is the number of agents,
A = A
1
× ... × A
n
the set of joint actions, Z is the set
of local observations, and O(s
t
, i) is the observation
function for agent i with 1 ≤ i ≤ n. R = R
1
× ... × R
n
is the joint reward function, with R
i
(s
t
, a
t,i
) being the
individual reward of agent i.
In MARL, each agent i has to find a local policy π
i
which is optimal w.r.t. the policies of the other agents.
If the other agents change their behavior then agent
i also need to adapt, since its previous policy might
have become suboptimal. The simplest approach to
MARL is to use single-agent RL algorithms like Q-
Learning and scale them up to multiple agents (Tan,
1993; Leibo et al., 2017). In homogeneous settings,
the policies can be shared by effectively learning only
one local policy π
i
and replicate the learned policy
to all agents. This can accelerate the learning pro-
cess as experience can be shared during training (Tan,
1993; Foerster et al., 2016). While many other ap-
proaches to MARL in games exist which incorporate
global information into the training process (Foerster
et al., 2016; Lowe et al., 2017; Foerster et al., 2018;
Rashid et al., 2018), we focus on the simple case of
applying single-agent RL to games with policy shar-
ing.
Nash Equilibria in Multi-Agent Swarms
235

2.3 Swarm Intelligence
One of the most common approaches for generating
artificial flocking behavior is the so called “Boids”
approach proposed by (Reynolds, 1987). Reynolds
proposed three basic steering rules which only require
local knowledge of an individual about other individ-
uals within its view radius. The rules are:
• Alignment: Steer towards the average heading di-
rection of nearby individuals
• Cohesion: Steer towards the average position
(center of mass) of visible individuals
• Separation: Steer in order to keep a minimum
distance to nearby individuals (to avoid collisions
and crowding)
If each individual follows these rules, naturally ap-
pearing swarm formations can be observed. Imple-
menting the rules can be done by expressing them as
forces that act upon an individual. They can be ex-
tended in order to repel from an enemy or obstacles
respectively be attracted by food, for example.
Hahn et al. (Hahn et al., 2019) introduced SELF-
ish, a Multi-Agent system in which multiple homoge-
neous agents (independently from each other) try to
survive as long as possible while a predator is chasing
them. The predator might get distracted by multiple
preys in its vicinity. Hahn et al. showed that agents
trained using Multi-Agent reinforcement learning re-
alize to exploit this property by forming a swarm in
order to increase their survival chances. This swarm-
ing behavior is extensively examined and compared
to other swarming/flocking algorithms (for example
the “Boids” approach). Furthermore (Hahn et al.,
2019) measured the survival time of the agents and
compared it to other policies, among others a hand
crafted policy called TurnAway. By following the
TurnAway strategy, agents turn in the opposite direc-
tion the predator and flee without considering other
agents or obstacles. Hahn et al. ended their paper
with a hypothesis of Nash equilibria in Multi-Agent
swarms. This hypothesis will be picked up and ex-
tended in this work.
¨
Ozg
¨
uler and Yıldız (
¨
Ozg
¨
uler and Yıldız, 2013)
also investigated Nash equilibria in Multi-Agent
swarms. To do so, they modeled the foraging pro-
cess of multiple agents as a non-cooperative N-player
game. They assumed that each agent wants to min-
imize its individual total effort in a time interval by
controlling its velocity. By establishing a nonlin-
ear differential equation in terms of positions of the
agents and solving this equation they show that the
game has a Nash equilibrium.
2.4 Nash Equilibria
Game theory considers strategic interactions within a
group of individuals. In doing so, the actions of each
individual affect the outcome and the individuals are
aware of this fact. In addition, the participating indi-
viduals are considered rational. This means, that they
have clearly defined goals within the possible out-
comes of the interactions and that they implement the
best available strategy to pursue their goals. Usually,
the rules of the game and rationality are well known.
Basically, two different forms of representation
exist: the normal form is used when the players
choose one strategy without knowing the others’
choices. The extensive form is used when some play-
ers know what other players have done while play-
ing. In many settings, no communication between the
players is possible or desired, which is why in the
following only the normal form game (also: strate-
gic form game) is described. A normal form game
G = (N, {A
i
}
i∈N
, {u
i
}
i∈N
) consists of a set of play-
ers N, a set of actions with A
i
for each player i and a
payoff function u
i
: A → R for each player i. The ac-
tion profile a = (a
1
, ..., a
n
) is a collection of actions,
one for each player, also called strategy profile. a
−i
is a strategy profile without the action of player i. All
possible collections of actions are also called space of
action profiles A = (a
1
, ..., a
n
) : a
i
∈ A
i
, i = 1, ..., n.
A Nash Equilibrium (NE) is a strategy profile so
that each strategy is a best response to all other strate-
gies. A best response is the reaction to an action that
maximizes the payoff.
Expressed in formal terms, a result a
∗
=
(a
∗
1
, ..., a
∗
n
) is a Nash Equilibrium, if for every player i
the following holds: u
i
(a
∗
i
, a
∗
−i
) ≥ u
i
(a
i
, a
∗
−i
) ∀a
i
∈ A
i
.
The most interesting feature about NEs is that they
are self-assertive: no player has an incentive to devi-
ate unilaterally.
Regarding the swarm environment in this pa-
per, the self interested agents correspond to the
non-cooperative players of a normal form game.
SELFish
DQN
and TurnAway, which will be explained
in the Section 3 match to the possible set of actions,
a player, respectively agent, can choose from. The re-
ward of a agent, which correlates to the survival time
of an individual corresponds to the payoff of a certain
strategy profile. With defining such a swarm environ-
ment as a normal form game it is possible to find NEs
with a common Nash solver. In this work the popular
Gambit Solver was used (McKelvey et al., 2016).
ICAART 2020 - 12th International Conference on Agents and Artificial Intelligence
236
2.5 Social Dilemmas in Multi-Agent
Systems
In many Multi-Agent Systems, agents need to co-
operate in order to maximize their own utilities.
In (de Cote et al., 2006), the authors analyze Multi-
Agent social dilemmas in which RL algorithms are
confronted with Nash Equilibria forming a rational
optimal solution on the short term but not being op-
timal in repeated interaction. They propose heuris-
tic principles to improve cooperation and overcome
such one-shot Nash Equilibrium strategies. However,
their experiments are limited to a prisoner’s dilemma
scenario with three agents and four actions. More
recently, (Leibo et al., 2017) demonstrated that the
learned behavior of agents in Multi-Agent systems
changes as a function of environmental factors. They
experimentally show how conflicts can emerge from
competition over shared resources and how the se-
quential nature of real world social dilemmas affects
cooperation. While their environments’ complexity is
comparable to this paper’s, their experiments are lim-
ited to two agents. Nevertheless, the Nash Equilibria
suspected in (Hahn et al., 2019) and further examined
in this paper shares many characteristics with the se-
quential social dilemmas introduced in (Leibo et al.,
2017). The swarming behavior learned is a strategy
spanning over multiple actions and is experimentally
shown to not be the optimal strategy for the collective
of agents. But as swarming behavior clearly requires
some form of coordination, the analysis of whether or
not swarming may be considered a defect in the sense
of (Leibo et al., 2017) is left open for further analysis.
3 EXPERIMENTAL SETUP
3.1 SELFish Environment
We use the same environment as (Hahn et al., 2019)
and extend their experiments. The agents can roam
freely in a two-dimensional area (see Figure 1). The
area wraps around at the edges, meaning an agent that
leaves it on the right side will immediately re-enter it
from the left (same with top and bottom). Aside with
the trained agents the environment is inhabited by a
predator which pursues a predefined static policy. The
predator can sense prey agents within a certain radius
and chooses one randomly as target (and keeps this
target for a certain time). This means that the preda-
tor might be distracted by multiple agents in its prox-
imity, which implies that it might be beneficial for an
agent to be close to others, thus modeling one of the
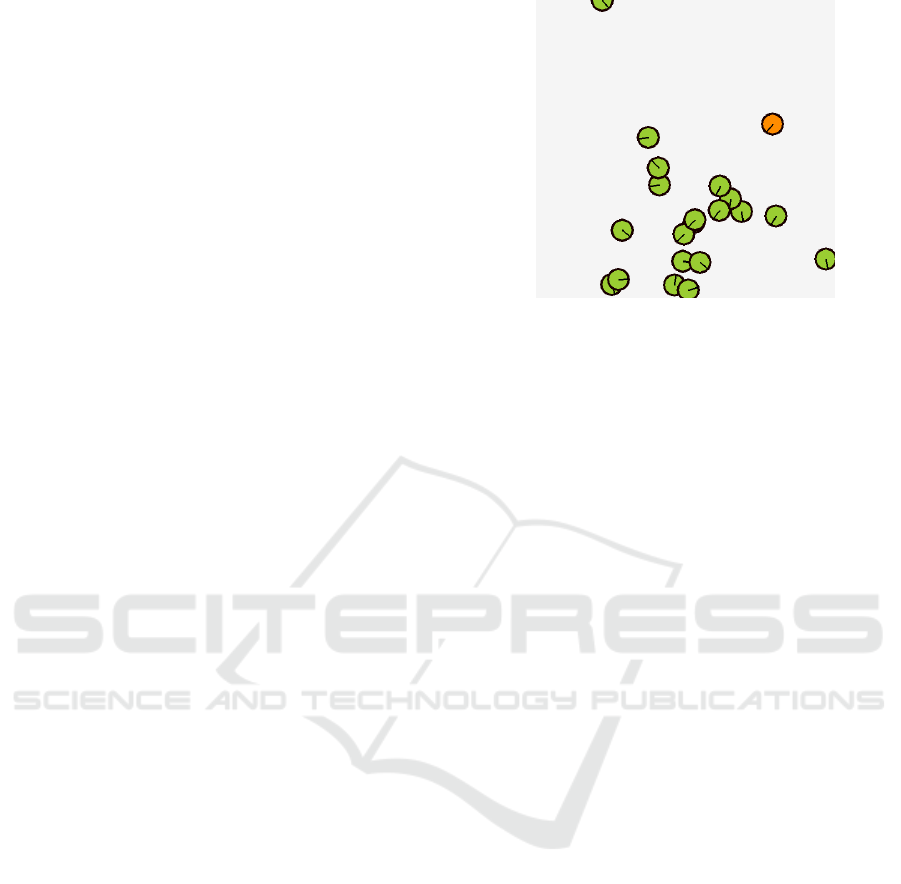
Figure 1: Example visualization of the environment show-
ing the agents in green and the predator in orange. The black
line shows the movement direction of the respective agent.
established benefits of joining a swarm. Both the prey
agents and the predator are embodied as circles with
a certain radius and a variable orientation (see Fig-
ure 1). A prey agent is considered caught when it
collides with the predator. It is then immediately re-
spawned in the area. The predator and the prey usu-
ally move at the same speed to encourage situations
in which a prey agent can escape from the predator
when it gets distracted. However, because of the torus
property of the area, this would allow a prey agent to
simply turn in the opposite direction of the predator
and move away without the possibility of the predator
ever catching up. That is why the predator increases
its speed every 80 steps over a duration of 20 steps.
The objective that is learned by the prey agents is
to survive as long as possible. This is reinforced with
a reward of +1 for each step survived and -1000 for
colliding with the predator. As soon as the learning
agent collides with the predator, the episode ends and
its knowledge is copied to all other agents.
As all agents move at a constant speed, the only
decision an agent has to make every step, is the
degree the agent wants to turn before it is moved
a certain unit in that direction. For DQN the ac-
tion space comprises of five discrete degree values
{−90
◦
, −45
◦
, 0
◦
, +45
◦
, +90
◦
}.
3.2 State Encoding
The state of the environment is only partially observ-
able for an agent. This facilitates the scalability of
the approach and represents autonomous agents or bi-
ological individuals in reality as it is unlikely that an
individual can sense the whole state of any physical
environment/world. It is also in accordance with other
related swarming algorithms like (Reynolds, 1987)
where an agent only considers a local neighborhood
Nash Equilibria in Multi-Agent Swarms
237
of other agents and adjusts its direction according to
rules of cohesion, alignment and separation
In SELFish (Hahn et al., 2019), every agent can at
most observe n other entities (i.e., other prey agents
or the predator) in its vicinity. As observation, ev-
ery agent a receives for all other agents e
i
, i ∈ [1, n],
within its observation the distance between a and e
i
,
the angle a would have to turn in order to face in the
direction of e
i
and the absolute orientation of e
i
in
space. The angle an agent has to turn in order to face
to another observed entity is pre-calculated in degrees
in the range of (−180
◦
, 180
◦
]. The absolute orienta-
tion of an entity in space is measured in degrees in
the range of [0
◦
, 360
◦
). 0
◦
corresponds to facing east-
wards, measuring the angle counter-clockwise.
Every agent receives the previously explained
measurements as observation for the predator, itself
and for a certain number of n neighboring agents, in
which the n neighbors are ordered by their distance.
The measurements are flattened into a vector before
they are handed to the agents. Furthermore the dis-
tance is divided by the area width and the direction
and orientation are normalized to the interval [0, 1].
3.3 Training
Training is performed using the Keras-RL (Plap-
pert, 2016) implementation of DQN. As DQN is in-
tended for single agent use, the training of the multi-
ple homogeneous agents is executed as proposed by
(Egorov, 2016): Only one agent is actually trained
and its policy is copied to the others after the end of
an episode. An episode ends if the learning agent is
caught by the predator or after 10, 000 steps were ex-
ecuted.
During the training the edge lengths of the area
always are 40 by 40 pixels. However, the agents as
well as the predator can take every real valued po-
sition in [0, 40] × [0, 40]. The agents and the preda-
tor have the size of a circle with radius 1, meaning a
collision (catch) occurs at a distance below 2. Please
note that no other collisions are considered (between
agents and other agents or agents and walls). Also
there are no obstacles in the area. During training
there are 10 agents in the environment.
In order to assess the quality of a training run, the
cumulative reward of the learning agent is measured.
According to the reward structure this essentially cor-
responds with the number of time steps the learning
agent survived.
Reproduced from (Hahn et al., 2019), the results
of training showed swarming behavior of the agents
in order to increase their survival chances, although
only 10 agents were present during training.
4 SCALING OF PARTIALLY
OBSERVABLE SCENARIOS
Because of the partial observability and the normal-
ization of the observation the learned policy of the
case with 10 agents can also be used in scenarios with
other parameters in regard to the number of agents
present or the size of the available area. This interre-
lationship will be further investigated in the following
section.
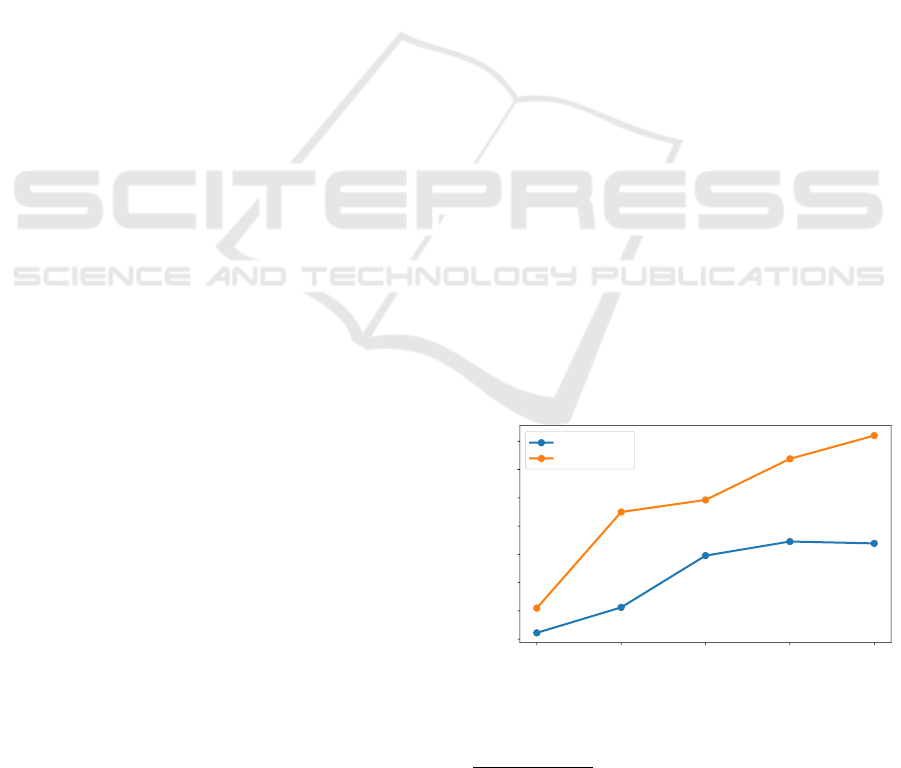
Figure 2 shows the average episode length, which
essentially corresponds to the average survival time of
a certain agent (i.e., the learning agent). An episode
ends if the learning agent is caught by the predator
or 10, 000 steps were made. As the learning agent
receives a +1 for every step and −1000 for being
caught, it is encouraged to survive as long as pos-
sible to maximize its accumulated reward. For the
static strategy of turning 180
◦
away from the preda-
tor without minding other agents (i.e., TurnAway) this
means that a certain agent is caught in order for an
episode to end.
1
. In Figure 2 the number of agents
is varied while the size of the area remains the same
(40×40 pixels). Please note that although the number
of agents is varied, the policy of SELFish
DQN
, which
was learned with 10 agents in the environment, stays
the same. One can see that the performance of the
policy, despite the fact that the environment settings
are modified, does not collapse. The increase of the
average episode length for higher number of agents
comes from the circumstance that, with more agents
in the environment, the probability decreases that any
particular agent is caught. The measurements for Tur-
nAway are given for comparison. Nevertheless, Fig-
ure 2 shows that TurnAway performs better w.r.t. the
20 40 60 80 100
Number of agents
1000
1500
2000
2500
3000
3500
4000
4500
Avg. episode lenght
SELFish
DQN
TurnAway
Figure 2: Average episode length (survival time) of the
learning agent in accordance with (Hahn et al., 2019). The
policy SELFish
DQN
was trained with 10 agents on 40 × 40
pixels and it then used for larger numbers of agents.
1
For a short video showing an example of the policies
SELFish
DQN
and TurnAway please refer to https://youtu.
be/nYKamj9qjFM
ICAART 2020 - 12th International Conference on Agents and Artificial Intelligence
238
average episode length or the survival time of a partic-
ular agent, respectively. This raises the question, why
this policy was not found by reinforcement learning,
which will be further examined in the following sec-
tion.
5 NASH EQUILIBRIA IN
SWARMS
As previously mentioned, 10 homogeneous agents
were trained on a 40 × 40 pixel area, while only one
agent was actually trained using reinforcement learn-
ing and its policy was copied to all other agents after
each episode. An episode ended if the learning agent
was caught by the predator (i.e. collided with it) or
10, 000 steps were executed. The reward was struc-
tured in such a way that the learning agent was en-
couraged to stay alive as possible. The predator has
the property that it might get distracted by multiple
prey agents in its proximity. That is why the agents
learned to form swarms in order to increase their sur-
vival chances. Another strategy in which the agent
always turned in the opposite direction of the preda-
tor and fled without minding other agents was imple-
mented for comparison (TurnAway). Figure 2 showed
that the policy learned with 10 agents on a 40 × 40
pixel area can be used in settings with more agents
without breaking. But furthermore it showed that the
static policy of turning away performs better than the
learned policy in terms of survival time.
To further investigate this phenomenon we carry
out an experiment in which agents pursuing both poli-
cies (learned and static) are present. The results are
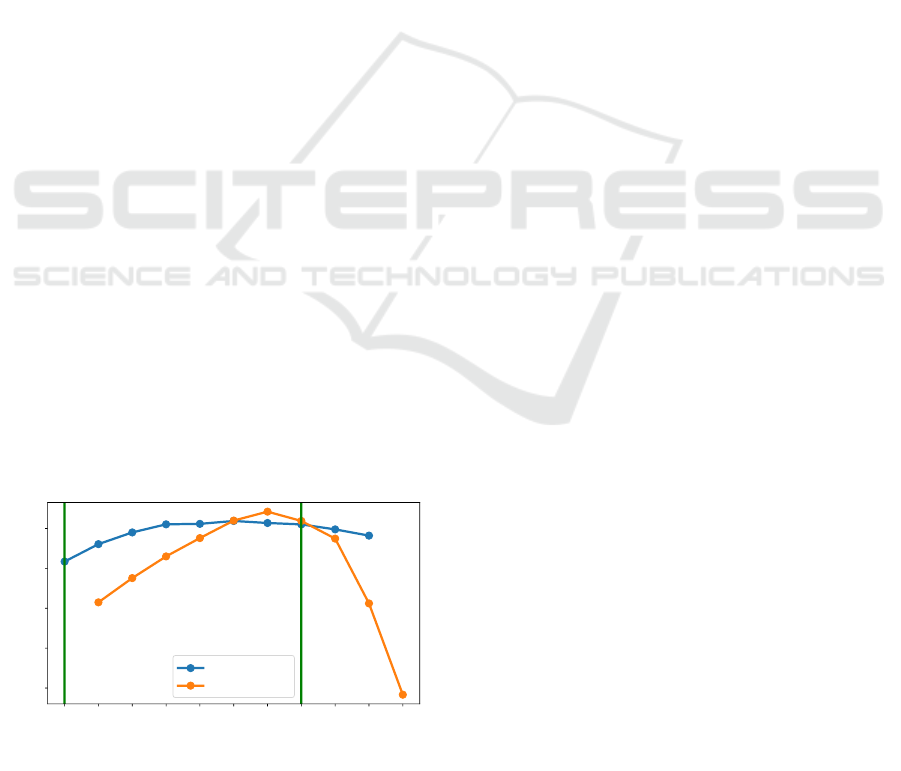
shown in Figure 3. Figure 3 shows a setup where
in an area of 40 × 40 pixels 10 agents are present.
These agents either use the learned SELFish
DQN
pol-
icy (trained with 10 homogeneous agents on 40 × 40
(0, 10)
(1, 9)
(2, 8)
(3, 7)
(4, 6)
(5, 5)
(6, 4)
(7, 3)
(8, 2)
(9, 1)
(10, 0)
(Number of TurnAway Agents, Number of SELFish
DQN
Agents)
200
300
400
500
600
Avg. survival time of an individual
SELFish
DQN
TurnAway
Figure 3: Performance of SELFish
DQN
(trained with 10
agents on 40×40 pixel) versus TurnAway on a 40×40 pixel
area. Nash equilibria marked with a green line.
pixel) or the static TurnAway policy. The agents
following SELFish
DQN
tend to form swarms as they
learned that this might increase their survival chances
(given the property of a distractible predator) while
TurnAway-agent do not care about others (except for
the predator). The abscissa illustrate the mixing pro-
portion of the agents with their particular type. For
example, the entry (6, 4) on the abscissa means that
there are 6 agents executing TurnAway and 4 agents
executing SELFish
DQN
in the setting. On the ordi-
nate the survival time (in steps) of individuals fol-
lowing either SELFish
DQN
or TurnAway is indicated
(averaged over individuals inside each policy type).
The experiment was carried out over 10 runs (with
different seeds), each with 10 episodes which lasted
100, 000 steps (with no other stopping criteria). This
resulted in thousand of caught agents in both pol-
icy groups whose survival time was then averaged.
Caught agents respawned at the most dense spot of the
swarm (determined with Kernel Density Estimation
(Phillips et al., 2006; Hahn et al., 2019)). This was
done because moving away from the swarm respec-
tively ignoring the action of other agents was of par-
ticular interest for this experiment and the swarm be-
havior should not be disturbed by respawning agents.
The experiment reveals multiple interesting in-
sights:
1. The performance of SELFish
DQN
w.r.t. the sur-
vival time in the setting it was actually trained on
is better than TurnAway. This means that the us-
age of a policy obtained in a partially observable
model in a setting with other parameters is possi-
ble but might not result in consistent performance
with the trained setting (cf. Figure 2).
2. Flocking behavior resulting from reinforcement
learning in this setting (multiple homogeneous
agents evading/distracting a predator) is a Nash
equilibrium (green line at (0, 10) in Figure 3).
This means that if there are 10 agents per-
forming SELFish
DQN
with a tendency to swarm-
ing/grouping (and zero performing TurnAway),
then no agent has an incentive to deviate from
this policy of “swarming” while all others keep
their strategy. We can see that moving out of the
swarm and ignoring the others (like TurnAway)
leads the agent onto the free space, where it is an
easy prey for the predator. We also confirmed this
intuition by calculating the Nash equilibria with
the Gambit software tools for game theory (McK-
elvey et al., 2016). This was done by consid-
ering the average survival time of an agent pur-
suing a certain strategy as payoff in a normal-
form game. This means, for example, that if 9
agents perform SELFish
DQN
, the payoff of ev-
Nash Equilibria in Multi-Agent Swarms
239
ery agent following this strategy is 561 while the
one agent performing TurnAway has a payoff of
415. Gambit also revealed a less obvious Nash
equilibrium at (7, 3). At this point an agent per-
forming TurnAway (payoff 619) has no incentive
to switch its strategy to SELFish
DQN
(payoff 614
of SELFish
DQN
at point (6, 4). In addition, an
agent performing SELFish
DQN
(payoff 610) has
no incentive to switch to TurnAway (payoff 574
at (8, 2)) while the other agents keep their strat-
egy (see Table 1).
3. Policies in Multi-Agent scenarios produced with
DQN and the method proposed by (Egorov, 2016)
can only take the outer points (0, 10) and (10, 0)
as all agents perform the same policy. In the ex-
ample of an area size of 40 × 40 pixels, (0, 10) is
surely better while a ratio of (6, 4) is the best mix-
ture of both strategies (although) still not the best
performance achievable in this setting.
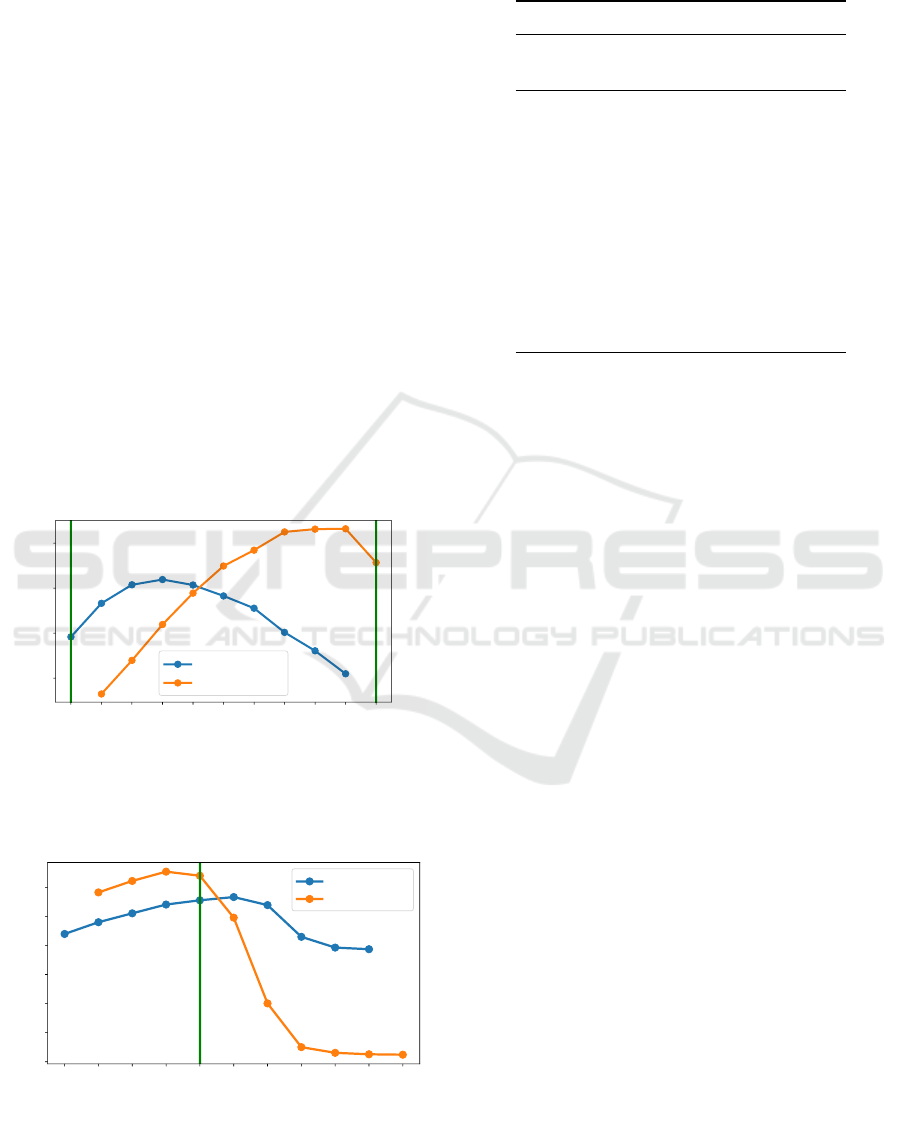
To further substantiate our results, we repeat this
experiment in area sizes of 80 ×80 and 20×20 pixels.
The results can be seen in Figure 4 and 5. They show
that the results vary for different area sizes, like pure
(0, 10)
(1, 9)
(2, 8)
(3, 7)
(4, 6)
(5, 5)
(6, 4)
(7, 3)
(8, 2)
(9, 1)
(10, 0)
(Number of TurnAway Agents, Number of SELFish
DQN
Agents)
1000
1200
1400
1600
Avg. survival time of an individual
SELFish
DQN
TurnAway
Figure 4: Performance of SELFish
DQN
(trained with 10
agents on 40×40 pixel) versus TurnAway on a 80×80 pixel
area. Nash equilibria marked with a green line.
(0, 10)
(1, 9)
(2, 8)
(3, 7)
(4, 6)
(5, 5)
(6, 4)
(7, 3)
(8, 2)
(9, 1)
(10, 0)
(Number of TurnAway Agents, Number of SELFish
DQN
Agents)
0
25
50
75
100
125
150
Avg. survival time of an individual
SELFish
DQN
TurnAway
Figure 5: Performance of SELFish
DQN
(trained with 10
agents on 40×40 pixel) versus TurnAway on a 20×20 pixel
area. Nash equilibria marked with a green line.
Table 1: Survival time/payoff of 10 agents with different
policies in an area of 40 × 40 pixels.
Number of agents Survival time
Turn SELFish Turn SELFish
Away
DQN
Away
DQN
0 10 nan 517
1 9 415 561
2 8 476 590
3 7 530 611
4 6 576 612
5 5 620 619
6 4 642 614
7 3 619 610
8 2 575 598
9 1 412 582
10 0 183 nan
TurnAway clearly outperforming pure SELFish
DQN
in the 80 × 80 pixel case and different Nash equilibria
like (4, 6) in the 20 × 20 pixel case.
6 CONCLUSION
In this paper we further examine the experimental
setup of (Hahn et al., 2019). Hahn et al. trained mul-
tiple homogeneous agents to evade a predator for as
long as possible. The predator in that setting follows
a static pre-defined policy and can be distracted by
multiple possible preys in its vicinity. This modelled
one of the benefits of forming a swarm and the agents
learned to exploit this circumstance accordingly.
Expanding this setup, we showed that policies ob-
tained through reinforcement learning in partially ob-
servable scenarios can be used in other settings with-
out a collapse of the performance, although consistent
performance (compared to the setting the policy was
actually trained on) cannot be guaranteed. Further-
more we showed that the swarm resulting from Multi-
Agent reinforcement learning in a predator/prey sce-
nario has a Nash equilibrium, i.e., that there are sce-
narios were specific swarm configurations are stable
(assuming rational agents) but still suboptimal. We
analyzed this effect in dependance of the area size.
We concluded swarming or not swarming can be for-
mulated as a social dilemma in some settings.
This sheds some light on the reasons why swarms
emerge. The introduction started by listing benefits
observed from biological swarms. The presented re-
search, however, might list another reason for the for-
mation of swarms: social pressure. The existence
of a swarm of substantial size may actively impede
ICAART 2020 - 12th International Conference on Agents and Artificial Intelligence
240

the survival of non-swarming individuals, thus urging
them to join the swarm even when it is suboptimal
to all individuals’ survival. Note that this affects ra-
tional agents, i.e., swarm participants that act locally
optimal at every single one of their decisions.
Interestingly, the phenomenon of pressure aris-
ing from lack of communication and control struc-
tures has been observed in natural evolution as
well (Dawkins, 1976). Thus, swarms can (un-
der certain conditions) also be interpreted as self-
perpetuating, which means that they should be han-
dled with additional care when employing them
in practical applications. Self-perpetuating swarms
might introduce additional targets for emergent be-
havior that affect the system designer’s intended pur-
pose. It is up to future research to examine the inter-
play between using such emergent behavior and con-
trolling it to employ useful swarm applications.
REFERENCES
Bellman, R. (1957). Dynamic Programming. Princeton
University Press, Princeton, NJ, USA, 1 edition.
Brambilla, M., Ferrante, E., Birattari, M., and Dorigo, M.
(2013). Swarm robotics: a review from the swarm
engineering perspective. Swarm Intelligence, 7(1):1–
41.
Christensen, A. L., Oliveira, S., Postolache, O., De Oliveira,
M. J., Sargento, S., Santana, P., Nunes, L., Velez, F. J.,
Sebasti
˜
ao, P., Costa, V., et al. (2015). Design of com-
munication and control for swarms of aquatic surface
drones. In ICAART (2), pages 548–555.
Dawkins, R. (1976). The Selfish Gene. Oxford University
Press, Oxford, UK.
de Cote, E. M., Lazaric, A., and Restelli, M. (2006). Learn-
ing to cooperate in multi-agent social dilemmas. In
Proceedings of the Fifth International Joint Confer-
ence on Autonomous Agents and Multiagent Systems,
AAMAS ’06, pages 783–785, New York, NY, USA.
ACM.
Egorov, M. (2016). Multi-agent deep reinforcement learn-
ing. CS231n: Convolutional Neural Networks for Vi-
sual Recognition.
Foerster, J., Assael, I. A., de Freitas, N., and Whiteson, S.
(2016). Learning to communicate with deep multi-
agent reinforcement learning. In Advances in Neural
Information Processing Systems, pages 2137–2145.
Foerster, J. N., Farquhar, G., Afouras, T., Nardelli, N., and
Whiteson, S. (2018). Counterfactual multi-agent pol-
icy gradients. In Thirty-Second AAAI Conference on
Artificial Intelligence.
Hahn, C., Phan, T., Gabor, T., Belzner, L., and Linnhoff-
Popien, C. (2019). Emergent escape-based flock-
ing behavior using multi-agent reinforcement learn-
ing. The 2019 Conference on Artificial Life, (31):598–
605.
Hausknecht, M. and Stone, P. (2015). Deep recurrent q-
learning for partially observable mdps. In 2015 AAAI
Fall Symposium Series.
Howard, R. A. (1961). Dynamic Programming and Markov
Processes. The MIT Press.
Leibo, J. Z., Zambaldi, V., Lanctot, M., Marecki, J., and
Graepel, T. (2017). Multi-agent reinforcement learn-
ing in sequential social dilemmas. In Proceedings of
the 16th Conference on Autonomous Agents and Mul-
tiAgent Systems, pages 464–473. International Foun-
dation for Autonomous Agents and Multiagent Sys-
tems.
Lowe, R., Wu, Y., Tamar, A., Harb, J., Abbeel, O. P.,
and Mordatch, I. (2017). Multi-agent actor-critic
for mixed cooperative-competitive environments. In
Advances in Neural Information Processing Systems,
pages 6379–6390.
McKelvey, R. D., McLennan, A. M., and Turocy, T. L.
(2016). Gambit: Software tools for game theory.
Mnih, V., Kavukcuoglu, K., Silver, D., Rusu, A. A., Veness,
J., Bellemare, M. G., Graves, A., Riedmiller, M., Fid-
jeland, A. K., Ostrovski, G., et al. (2015). Human-
level control through deep reinforcement learning.
Nature, 518(7540):529–533.
Morihiro, K., Nishimura, H., Isokawa, T., and Matsui,
N. (2008). Learning grouping and anti-predator be-
haviors for multi-agent systems. In Int’l Conf. on
Knowledge-Based and Intelligent Information and
Engineering Systems. Springer.
¨
Ozg
¨
uler, A. B. and Yıldız, A. (2013). Foraging swarms as
nash equilibria of dynamic games. IEEE transactions
on cybernetics, 44(6):979–987.
Phillips, S. J., Anderson, R. P., and Schapire, R. E. (2006).
Maximum entropy modeling of species geographic
distributions. Ecological modelling, 190(3-4).
Pinciroli, C. and Beltrame, G. (2016). Swarm-oriented pro-
gramming of distributed robot networks. Computer,
49(12):32–41.
Plappert, M. (2016). keras-rl. https://github.com/keras-
rl/keras-rl.
Puterman, M. L. (2014). Markov decision processes: dis-
crete stochastic dynamic programming. John Wiley &
Sons.
Rashid, T., Samvelyan, M., Witt, C. S., Farquhar, G., Foer-
ster, J., and Whiteson, S. (2018). Qmix: Monotonic
value function factorisation for deep multi-agent rein-
forcement learning. In International Conference on
Machine Learning, pages 4292–4301.
Reynolds, C. W. (1987). Flocks, herds and schools: A dis-
tributed behavioral model. In ACM SIGGRAPH com-
puter graphics, volume 21. ACM.
Tan, M. (1993). Multi-agent reinforcement learning: in-
dependent versus cooperative agents. In Proceed-
ings of the Tenth International Conference on Interna-
tional Conference on Machine Learning, pages 330–
337. Morgan Kaufmann Publishers Inc.
Watkins, C. J. C. H. (1989). Learning from Delayed Re-
wards. PhD thesis, King’s College, Cambridge, UK.
Nash Equilibria in Multi-Agent Swarms
241
