
EVOLUTIONARY SEARCH IN LETHAL ENVIRONMENTS
Richard Allmendinger and Joshua Knowles
School of Computer Science, University of Manchester, Manchester, U.K.
Keywords:
Evolutionary computation, Closed-loop optimization, Mutational robustness, Embodied evolution, Evolvable
hardware, Evolvability, NKα landscapes.
Abstract:
In Natural evolution, a mutation may be lethal, causing an abrupt end to an evolving lineage. This fact has a
tendency to cause evolution to “prefer” mutationally robust solutions (which can in turn slow innovation), an
effect that has been studied previously, especially in the context of evolution on neutral plateaux. Here, we
tackle related issues but from the perspective of a practical optimization scenario. We wish to evolve a finite
population of entities quickly (i.e. improve them), but when a lethal solution (modelled here as one below a
certain fitness threshold) is evaluated, it is immediately removed from the population and the population size
is reduced by one. This models certain closed-loop evolution scenarios that may be encountered, for example,
when evolving nano-technologies or autonomous robots. We motivate this scenario, and find that evolutionary
search performs best in a lethal environment when limiting randomness in the solution generation process, e.g.
by using elitism, above-average selection pressure, a less random mutating operator, and no or little crossover.
For NKα landscapes, these strategies turn out to be particularly important on rugged and non-homogeneous
landscapes (i.e. for large K and α).
1 INTRODUCTION
In this paper, we consider the use of evolutionary al-
gorithms (EAs) for optimization in a setting where the
notions of an individual and of a population size, are
slightly different from standard, which forces us to re-
consider how to configure an EA appropriately. The
general setting is closed-loop optimization (Klock-
gether and Schwefel, 1970; Harvey et al., 1996;
Rechenberg, 2000; Shir et al., 2007; Calzolari et al.,
2008; Allmendinger and Knowles, 2011), which is an
area in resurgence, and likely to expand further in fu-
ture. In these type of problems, candidate solutions
(genotypes) to an optimization problem are planned
on a computer but their phenotypes are realized or
prototyped using experiments and/or real hardware;
the process of measuring the fitness (or quality) of
the phenotype may also involve conducting an exper-
iment and/or the use of hardware. There are several
ways in which this kind of setup may cause difficul-
ties, and there has been recent work (Allmendinger
and Knowles, 2011) considering some aspects of
closed-loop evolution related to dynamic constraints
and the dynamic availability of resources. Here, our
concern is for a particular sort of closed-loop setting
where the hardware on which individuals are tested
are reconfigurable, destructible and non-replaceable.
Consider the following scenario. We are devel-
oping, by evolution, the control software for an au-
tonomous endoscopic robot (similar to (Moglia et al.,
2007; Ciuti et al., 2010)), which is ultimately intended
to be swallowed by a patient in capsule form, and
then used within the body for screening, diagnosis and
therapeutic procedures. Before robotic capsules are
used on humans, however, their reliability and effec-
tiveness typically needs to be first validated through
in vivo animal trials. Imagine we have available µ
prototypes of a robotic capsule, and we can “radio in”
new control software to each of these individual cap-
sules and evaluate them. However, our control soft-
ware may cause a capsule to malfunction in a lethal
(for it) way, in which case it is no longer available
(and falls out of the evolving population). In such
a scenario, our population size for the remainder of
the evolution is now at most µ− 1, in terms of how
many pieces of software can be tested each genera-
tion. If we continue to explore too aggressively, we
may upload other pieces of software that cause the
loss of individuals, which will cost evolution in terms
of the extent of parallelism (and hence efficiency in
time) we enjoy, and also perhaps in the loss of expen-
sive hardware. So whilst we wish to innovate by test-
63
Allmendinger R. and Knowles J..
EVOLUTIONARY SEARCH IN LETHAL ENVIRONMENTS.
DOI: 10.5220/0003673000630072
In Proceedings of the International Conference on Evolutionary Computation Theory and Applications (ECTA-2011), pages 63-72
ISBN: 978-989-8425-83-6
Copyright
c
2011 SCITEPRESS (Science and Technology Publications, Lda.)

ing new control policies, this must be balanced care-
fully against considerations of maintaining a reason-
able population size.
This setting may seem far-fetched to some read-
ers. However, as we see evolution being applied more
and more at the boundary between a full embodied
type of evolution (Watson et al., 2002; Zykov et al.,
2005) (where the hardware individuals may repro-
duce or exchange information) and a fully in silico
simulation-only approach, especially as EAs are taken
up in the experimental sciences more and more, it is
likely that variants of the above scenario will emerge.
It need not be robotic capsules; it could equally be
nano-machines or drone planes we are dealing with;
it could be robot swarms, or software released on the
Internet, or anything that is reconfigurable remotely
in some way, and also destructible.
Clearly, the principle of “survival of the fittest” is
going to be somewhat dampened by the constraint that
we wish all designs to survive. Indeed, no innovation
at all will be possible if we enforce the evaluation of
only safe designs, given the assumption that any un-
seen design could be a lethal one. So, as is usual with
evolution, we should expect that some rate of loss of
individuals is going to be desirable, but set against
this is the fact that they cannot be replaced, and hence
population size and parallelism will be compromised.
So, the question is, how should one control the explo-
ration/exploitation tradeoff in this optimization setup.
In more Natural settings than we consider here,
the notion of robustness and the tendency of evolu-
tion to build in mutationalrobustness for free has been
previously studied (Bullock, 2003; Schonfeld, 2007).
Bullock’s work in particular shows that certain in-
built biases toward mutational robustness can retard
innovation (our primary concern here). He proposed
methods that avoid the bias and hence explore neutral
plateaux more rapidly. This work has a different pur-
pose to ours, and involves different assumptions, but
there are also obvious parallels which we hope will be
apparent from our results.
Our more practical investigation of the effects on
optimization performance of lethal mutations looks
at some of the simple EA configuration parameters:
degree-of-elitism, population size, selection pressure,
mutation mode and strength, and crossover rate. We
also vary aspects of the fitness landscape we are opti-
mizing to observe which landscape topologies pose a
particular challenge when optimizing in a lethal envi-
ronment.
The rest of the paper is organized as follows. The
next section describes the experimental setup includ-
ing the search algorithms considered (Section 2.1)
and the family of binary test problems (NKα land-
scapes) we use to analyze the impact of lethal solu-
tions (Section 2.2). Experimental results are given in
Section 3; we draw conclusions in Section 4. Finally,
Section 5 outlines possible directions for future work.
2 EXPERIMENTAL SETUP
This section describes the family of binary test func-
tions f we consider, the search algorithms for which
we investigate the impact of a lethal optimization sce-
nario, and the parameter settings as used in the subse-
quent experimental analysis.
2.1 Search Algorithms
We consider three types of search algorithms in this
work: a tournament selection based genetic algorithm
(TGA), a modified version of it which we are call-
ing RBS (standing for reproduction of best solutions),
and a population of stochastic hill-climbers (PHC).
Similar types of algorithms have been considered in
previous studies related to mutational robustness (see
e.g. (Bullock, 2003; Schonfeld, 2007)).
Before we describe each algorithm, we first set out
the procedures common to all of them. These are the
generation process of the initial population of candi-
date solutions, the setting of the fitness threshold be-
low which solutions are deemed lethal, the mutation
operators, and the handling of duplicate solutions.
Regarding the initialization, our aim is to simulate
the scenario where the initial population, whose size
we denote by µ
0
, consists of evolving entities (e.g.
robotic capsules, robot swarms or software) that are of
a certain (high or state-of-the-art) quality. We achieve
this by first generating a sample set of S random and
non-identical candidate solutions, and then selecting
the fittest µ
0
solutions from this set to form the initial
population. The lethal fitness threshold f
LFT
is set to
the fitness value of the qth fittest solution of the sam-
ple set. This threshold is kept constant throughout an
algorithmic run.
For mutation, we will investigate two modes (sim-
ilar to (Barnett, 2001)): (i) Poisson mutation, where
each bit is flipped independently with a mutation rate
of p
m
, and (ii) constant mutation, where exactly d
randomly selected bits are flipped. Furthermore, all
search algorithms ensure that a solution is not evalu-
ated multiple times. That is, all evaluated solutions
are cached and compared against a new solution (off-
spring) before performing an evaluation; preliminary
experimentation showed that avoiding duplication im-
proves the performance significantly. If a solution
has been evaluated previously, then the GA iteratively
ECTA 2011 - International Conference on Evolutionary Computation Theory and Applications
64

Algorithm 1: Tournament selection based GA (TGA).
Require: f (objective function), G (maximal number of
generations), µ
0
(initial parent population size), λ
0
(ini-
tial offspring population size), L (maximal number of re-
generation trials)
g = 0 (generation counter), Pop =
/
0 (current population),
OffPop =
/
0 (offspring population), AllEvalSols =
/
0 (set
of solutions evaluated so far), trials = 0 (regeneration tri-
als counter), counter = 0 (auxiliary variable indicating
the number of solutions evaluated during a generation)
Initialize Pop and set lethal fitness threshold f
LFT
; copy
all solutions of Pop also to AllEvalSols, and set µ
g
=
µ
0
,λ
g
= λ
0
while g < G ∧ µ
g
> 0 do
OffPop =
/
0,trials = 0,counter = 0
Set mutation rate to initial value
while counter < λ
g
do
Generate two offspring ~x
(1)
and ~x
(2)
by selecting
two parents from Pop, and then recombining and
mutating them
for i = 1 to 2 do
if~x
(i)
/∈ AllEvalSols ∧ counter < λ
g
then
Evaluate~x
(i)
using f , counter++, trials = 0
AllEvalSols = AllEvalSols∪~x
(i)
if f(~x
(i)
) ≥ f
LFT
then
OffPop = OffPop∪~x
(i)
else trials++
if trials = L then
Depending on the mutation operator, reset
mutation rate to p
m
= p
m
+ 0.5/N (Poisson
mutation) or d = d + 1 (constant mutation)
g++
Reset new population sizes to µ
g
= λ
g
= |OffPop|, and
form new population Pop by selecting the best µ
g
so-
lutions from the union population of Pop∪ OffPop
generates new solutions (offspring solutions) until it
generates one that is evaluable, i.e. has not been eval-
uated previously, or until L trials have passed without
success. In the latter case, we reset the mutation rate
to p
m
= p
m
+0.5/N (N is the total number of bits) and
d = d+1 in the case of Poisson mutation and constant
mutation, respectively; note, due to its deterministic
nature, it is more likely that this reset step is applied
with constant mutation. For TGA and RBS we set
the mutation rates back to their initial values at the
beginning of each new generation; for PHC it makes
more sense to set the mutation rates back whenever
a hill-climber of the current population is considered
the first time for mutation (see pseudocode of PHC).
2.1.1 Tournament Selection based GA (TGA)
The algorithm uses tournament selection with re-
placement for parental selection (TS shall denote the
tournament size) and uniform crossover (Syswerda,
1989) (p
c
shall denote the crossover rate). Our TGA
uses a (µ+ λ)-ES reproduction scheme, an elitist ap-
proach, which performed significantly better than a
standard generational reproduction scheme in prelim-
inary experimentation, and which we believe is gen-
erally applicable in this domain. We set the number
of offspring solutions to be identical to the population
size (or the number of parent solutions), i.e. µ = λ.
As the population size may decrease during the opti-
mization process (due to evaluated lethal solutions),
in future, we will denote the population size at gener-
ation g (0 ≤ g ≤ G) by µ
g
= λ
g
with g = G being the
maximum number of generations. Clearly, the max-
imum number of generations can only be reached if
the population does not “die out” beforehand. Algo-
rithm 1 shows the pseudocode of TGA.
RBS is based on TGA and motivated by the
netcrawler (Barnett, 2001), an algorithm that repro-
duces only a single best solution at each generation.
RBS selects parents for reproduction exclusively and
at random from the set of solutions of the current pop-
ulation with the highest fitness. Hence, it can be seen
as a tournament selection based GA with TS = µ
g
and
where the tournament participants are selected with-
out replacement. Apart from the selection procedure,
RBS will use the same algorithmic setup as TGA.
2.1.2 Population of Hill-climbers (PHC)
This search algorithm maintains a population of
stochastic hill-climbers, which explore the landscape
independently of each other. Each hill-climber under-
goes mutation and, if the mutant or offspring is not
lethal, it replaces the original solution if it is at least
as fit. Algorithm 2 shows the pseudocode of PHC. In
essence, PHC with constant mutation is identical to
running a set of random-mutation hill-climbers (For-
rest and Mitchell, 1993) in parallel with the difference
that, as in the case of TGA, the mutation strength is
temporarily increased if a previously unseen solution
cannot be generated within L trials. PHC has also
similarities to the netcrawler (Barnett, 2001) and the
plateau crawler (Bullock, 2003). These approaches
have in common that they guarantee that each parent
potentially leaves a copy or a slightly modified ver-
sion of itself in the population. This ensures that off-
spring, parents, or other ancestors, are equally likely
to be selected as future parents. For fitness landscapes
featuring neutral networks, this feature can cause a
population to be unaffected by mutational robustness
as it allows the population to spend at each point on
a neutral network an equal amount of time (Hughes,
1995; Bullock, 2003).
EVOLUTIONARY SEARCH IN LETHAL ENVIRONMENTS
65

Algorithm 2: Population of hill-climbers (PHC).
Require: f (objective function), G (maximal number of
generations), µ
0
(initial parent population size), L (max-
imal number of regeneration trials)
g = 0 (generation counter), Pop =
/
0 (current population),
OffPop =
/
0 (offspring population), AllEvalSols =
/
0 (set
of solutions evaluated so far), trials = 0 (regeneration tri-
als counter), lethal = false (auxiliary boolean variable in-
dicating whether a hill-climber is lethal)
Initialize Pop and set lethal fitness threshold f
LFT
; copy
all solutions of Pop also to AllEvalSols, and set µ
g
= µ
0
while g < G ∧ µ
g
> 0 do
OffPop =
/
0,trials = 0
for i = 1 to µ
g
do
Set mutation rate to initial value, lethal = false
repeat
Mutate solution~x
i
of Pop to obtain mutant~x
′
i
if~x
′
i
/∈ AllEvalSols then
Evaluate~x
′
i
using f, trials = 0
AllEvalSols = AllEvalSols∪~x
′
i
if f(~x
′
i
) ≥ f
LFT
then
if f(~x
′
i
) ≥ f(~x
i
) then
OffPop = OffPop∪~x
′
i
else OffPop = OffPop∪~x
i
else lethal = true
else trials++
if trials = L then
Depending on the mutation operator, reset
mutation rate to p
m
= p
m
+ 0.5/N (Poisson
mutation) or d = d + 1 (constant mutation)
until~x
′
i
is evaluable ∨ lethal = true
g++
Reset new population size to µ
g
= |OffPop|, and form
new population Pop by copying all solutions from
OffPop to Pop
2.2 Test Functions f
Our aim in this study is to understand the effect of
lethal solutions on EA performance on real closed-
loop experimental problems (ultimately). Hence, it
might be considered ideal to use, for testing, some set
of real-world closed-loop optimization problems sub-
ject to lethal solutions, that is: real experimental prob-
lems featuring real resources that may potentially be
fatally damaged. That way we could see the effects of
EA design choices directly on a real-world problem
of interest. But even granting this to be an ideal ap-
proach, it would be very difficultto achieve in practice
due to the inherent cost of conducting closed-loop ex-
periments and the difficulty of repeating them to ob-
tain any statistical confidence in results seen. For this
reason, our study will use more familiar artificial test
problems augmented with the possibility of encoun-
tering lethal candidate solutions.
The family of test functions we consider here is a
Table 1: Default parameter settings of search algorithms.
Algorithm Parameter Setting
TGA,
PHC,
RBS
Parent population size µ
0
30
Constant mutation rate d 1
Regeneration trials L 1000
Number of generations G 200
Sampling size S 1000
Solution rank q for lethal
250
fitness threshold setting
TGA
Offspring population size λ
0
30
Tournament size TS
4
(Selection with replacement)
Crossover rate p
c
0.0
RBS
Offspring population size λ
0
30
Tournament size TS
µ
g
(Selection without replacement)
Crossover rate p
c
0.0
variant of NK landscapes called NKα land-
scapes (Hebbron et al., 2008). The use of this
test problem should help us to gain initial insights
into the effects of lethal solutions on fitness land-
scapes featuring various degrees of epistasis and
ruggedness.
2.2.1 NKα Landscapes
The general idea of the NKα model (Hebbron
et al., 2008) is to extend Kauffman’s original NK
model (Kauffman, 1989) to model epistatic net-
work topologies that are more realistic in mapping
the epistatic connectivity between genes in natural
genomes. The NKα model achieves this by affecting
the distribution of influences of genes in the network
in terms of their connectivity, through a preferential
attachment scheme. The model uses a parameter α
to control the positive feedback in the preferential at-
tachment so that larger α result in a more non-uniform
distribution of gene connectivity. There are three tun-
able parameters involvedin the generation of an NKα
landscape: the total number of variables N, the num-
ber of variables that interact epistatically at each of
the N loci, K, and the model parameter α that allows
us to specify how influential some variables may be
compared to others. As α increases, an increasing in-
fluence is given to a minority of variables, while, for
α = 0, the NKα model reduces to Kauffman’soriginal
NK model with neighbors being selected at random;
this model has already been used previously to ana-
lyze certain aspects of real-world closed-loop prob-
lems (see e.g. (Thompson, 1996)).
2.3 Parameter Settings
The experimental study will investigate different set-
tings of the parameters involved in the search algo-
ECTA 2011 - International Conference on Evolutionary Computation Theory and Applications
66
rithms. However, if not otherwise stated, we use the
default settings as given in Table 1 with constant mu-
tation being the default mutation operator. Remember
that RBS uses the same default parameter settings as
TGA with the difference that reproductive selection
is done with TS = µ
g
and tournament participants are
chosen without replacement.
For NKα landscapes, we analyze the impact of
lethal solutions using different settings of the neigh-
borhood size K and the model parameter α. We fix
the search space size to N = 50, which corresponds
to a typical search space size we have seen in related
closed-loop problems (e.g. a drug discovery problem
with a library of 30–60 drugs (Small et al., 2011)).
Any results shown are average results across 100
independent algorithm runs. We will use a differ-
ent randomly generated problem instance for each run
but, of course, the same instances for all algorithms.
Furthermore, to allow for a fair comparison, we also
use the same initial population and (hence) lethal fit-
ness threshold for all algorithms in any particular run.
3 EXPERIMENTAL RESULTS
Before we analyze the effect of lethal solutions on
evolutionary search, we first give some indication of
the properties of the NKα landscapes, and the effect
of varying K and α for (N=50) over the ranges used in
our experiments. For this, we follow the experimen-
tal methods employing adaptive walks already used
by (Hebbron et al., 2008). Starting from a randomly
generated solution, an adaptivewalk calculates the fit-
ness of all neighbors of the solution that can be gener-
ated with a single mutation step, and selects one of the
fitter neighbors at random to move to. If there is no fit-
ter neighbor, then the walk has reached a local optima
and terminates. We have performed 10000 adaptive
walks for different values of K and α, and recorded
the length of each walk and the genotype of the lo-
cal optima where each walk terminated. We have re-
peated this process 10 times for different values of K
and α using always a different randomly generated
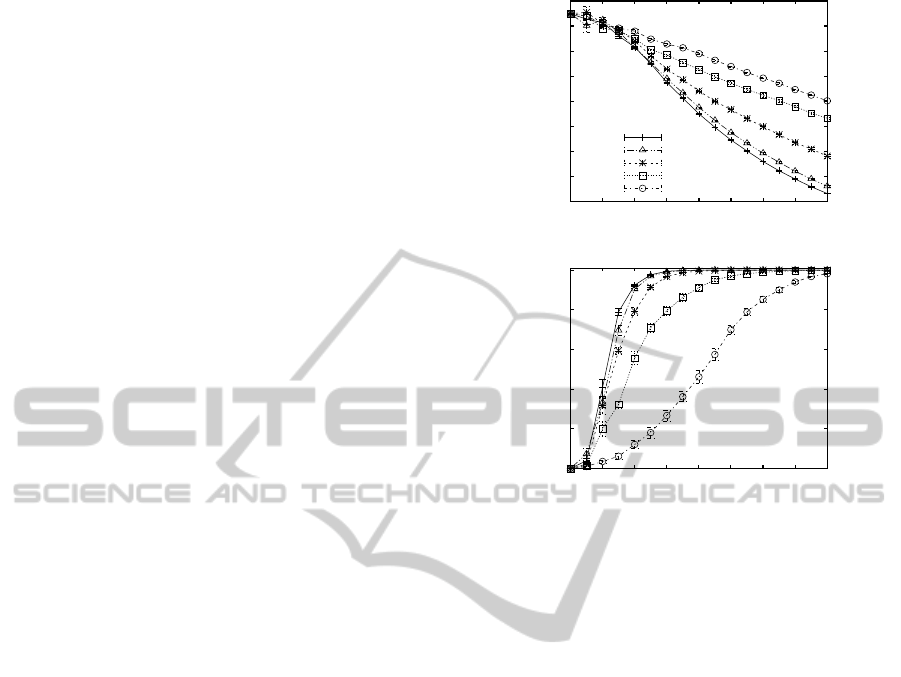
landscape. Figure 1 shows the average adaptive walk
length (top plot) and the average number of unique
(local) optimal solutions (bottom plot). In essence,
this figure shows that increasing K reduces the aver-
age length of an adaptive walk, i.e. the landscapes
becomes more rugged, and at the same time increases
the probability that the EA gets trapped at a new local
optimal solution. On the other hand, increasing α re-
duces the ruggedness of the fitness landscapes and de-
creases the number of unique local optimal solutions.
As is apparent from the figure, the parameters K and
10
12
14
16
18
20
22
24
26
0 2 4 6 8 10 12 14 16
Average adaptive walk length
#Neighbors K
α=0.0
α=0.5
α=1.0
α=1.5
α=2.0
0
2000
4000
6000
8000
10000
0 2 4 6 8 10 12 14 16
Average number of unique local
optimal solutions found
#Neighbors K
Figure 1: Plots showing the average length (top) and the av-
erage number of unique local optimal solutions found (bot-
tom) across 10 repetitions of 10000 adaptive walks for dif-
ferent neighborhood sizes K and model parameter values α.
α allow us to cover a reasonable mixture of fitness
landscapes exhibiting different topologies or levels of
ruggedness and epistasis.
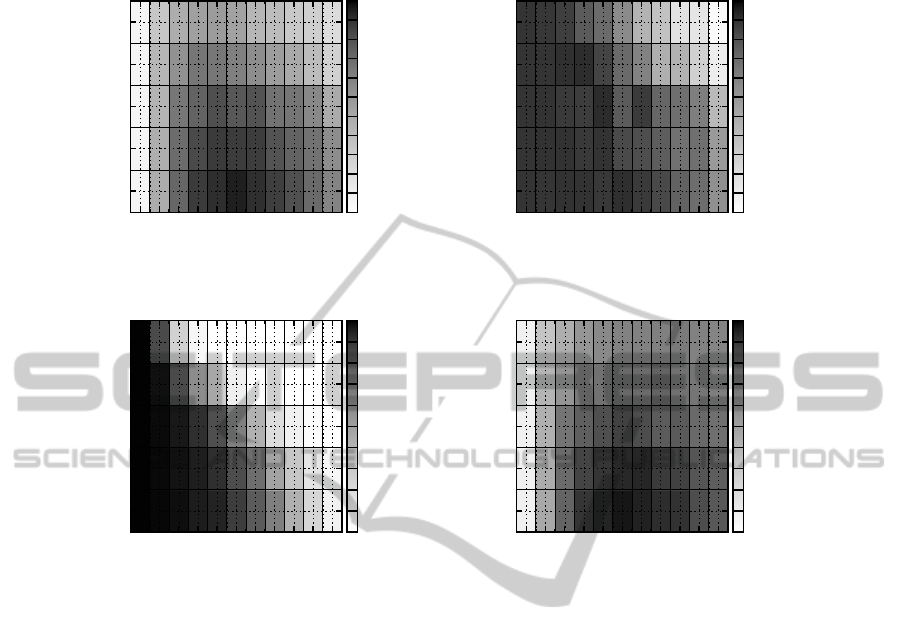
Figure 2 shows how a lethal optimization scenario
affects the best solution fitness found (top plots) and
the (remaining) population size (bottom left plot) for
TGA with a rather traditional setup (TS = 2, p
c
= 0.0
and Poisson mutation with p
m
= 1/N) as a function
of the parameters K and α; the bottom right plot in-
dicates the scope available for optimization after the
initialization of the population. From the top plots we
can see that the performance is most affected for fit-
ness landscapes that are rugged (large K) but contain
some structure in the sense that some solution bits are
more important than others (large α). As is apparent
from the bottom left plot, the reason for this pattern is
that the population size at the end of the optimization
decreases as K and/or α increase, i.e. lethal solutions
are here more likely to be encountered. In fact, for
large K and α, the optimization terminates on average
after only about 25 generations (out of 200). The rea-
son that the population size reduces for large K is that
the fitness landscape becomes more rugged, which in-
creases the risk of evaluating lethal solutions despite
them being located close to high-quality solutions.
Although an increase in α introduces more structure
into the fitness landscape, it has also the effect that so-
EVOLUTIONARY SEARCH IN LETHAL ENVIRONMENTS
67
TGA - Average best solution fitness
obtained in lethal environment
0 1 2 3 4 5 6 7 8 9 10
#Neighbors K
0
0.5
1
1.5
2
Model parameter α
0.66
0.67
0.68
0.69
0.7
0.71
0.72
0.73
0.74
0.75
0.76
0.77
TGA - Average absolute difference between
the best solution fitness found in
a lethal and non-lethal environment
0 1 2 3 4 5 6 7 8 9 10
#Neighbors K
0
0.5
1
1.5
2
Model parameter α
-0.05
-0.045
-0.04
-0.035
-0.03
-0.025
-0.02
-0.015
-0.01
-0.005
0
0.005
TGA - Survived proportion of the population
at generation G=200, or µ
G
/µ
0
0 1 2 3 4 5 6 7 8 9 10
#Neighbors K
0
0.5
1
1.5
2
Model parameter α
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
TGA - Average absolute difference between
the best solution fitness found in a non-lethal
environment and the lethal fitness threshold
0 1 2 3 4 5 6 7 8 9 10
#Neighbors K
0
0.5
1
1.5
2
Model parameter α
0.14
0.15
0.16
0.17
0.18
0.19
0.2
0.21
0.22
0.23
0.24
Figure 2: Plots showing the average best solution fitness found by TGA (using TS = 2, p
c
= 0.0 and Poisson mutation with
p
m
= 1/N) in a lethal environment (top left), the average absolute difference between this fitness and the fitness achieved
in a non-lethal environment, i.e. f
LFT
= 0 (top right), the average proportion of the population that survived at generation
G = 200, or µ
G
/µ
0
(bottom left), and the average absolute difference between the best solution fitness found and the lethal
fitness threshold in a non-lethal environment (bottom right), on NKα landscapes as a function of the neighborhood size K and
the model parameter α.
lutions which havesome of these important bits set in-
correctly are more likely to be poor. A plausible alter-
native explanation for the poor performance at large
α may be that the lethal fitness threshold is very close
to the best solution fitness that can be found, thus re-
ducing the scope of optimization possible and causing
many solutions to be lethal; the bottom right plot rules
this explanation out. Regarding the (remaining) pop-
ulation size, a greater selection pressure, i.e. a larger
tournament size, can help to maintain a large popula-
tion size for longer, particularly for large K and small
α. On the other hand, being more random in the solu-
tion generation process by using, for example, a larger
mutation and/or crossover rate, has the opposite ef-
fect, i.e. the population size decreases more quickly.
The effect on the (remaining) population size
translates into an impact on the average best solution
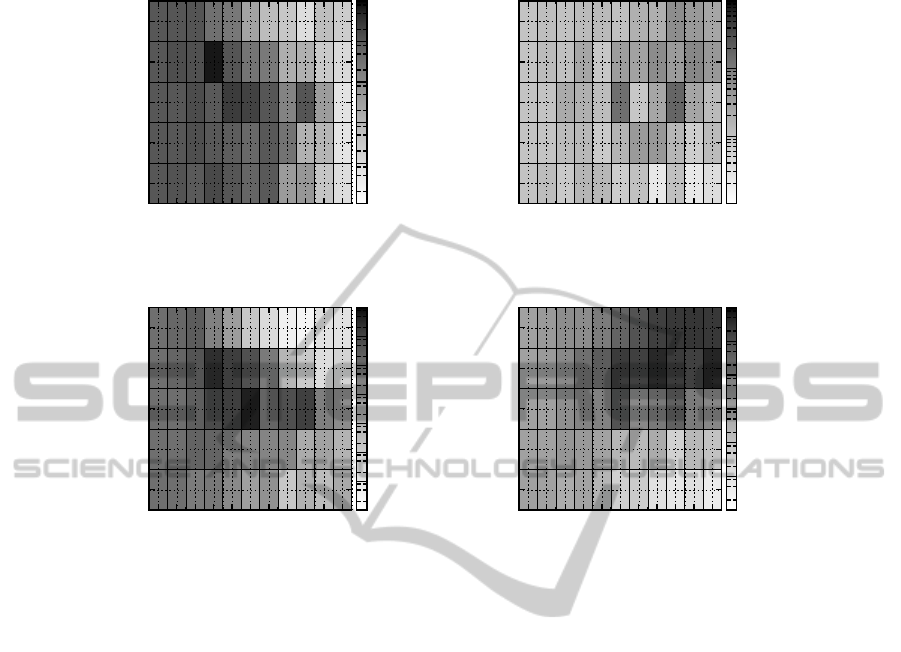
fitness. In fact, from Figure 3, we can see that, for
rugged landscapes with structure (i.e. in the range
K > 5, α > 1), TGA is outperformed by a GA with
a rather traditional setup and a PHC in a non-lethal
optimization scenario (right plots) but performs bet-
ter than both algorithms in a lethal optimization sce-
nario (left plots). In fact, as we will also see later, in
the range K > 5, α > 1, PHC performs best among all
the search algorithms considered in the non-lethal en-
vironment but worst in the lethal environment. The
poor performance of PHC in this range is due to the
fact that the hill-climbers in the population are search-
ing the fitness landscape independently of each other.
This may be an advantage in a non-lethal environment
because it can help to maintain diversityin the popula-
tion and prevent premature convergence. However, in
a lethal environment, uncontrolleddiversity is rather a
drawback as it increases the probability of generating
solutions with genotypes that are significantly differ-
ent from the ones in the population, which in turn in-
creases the probability of generating lethal solutions.
The less diverse and random optimization of TGA is
also the reason that it outperforms a GA with a more
ECTA 2011 - International Conference on Evolutionary Computation Theory and Applications
68
Ratio P(f(x) > f
TGA
)/P(f(x) > f
TGA,TS=2,p
c
=0.0,p
m
=1/N
),
lethal optimization
0 1 2 3 4 5 6 7 8 9 10
#Neighbors K
0
0.5
1
1.5
2
Model parameter α
10
-4
10
-3
10
-2
10
-1
10
0
10
1
Ratio P(f(x) > f
TGA
)/P(f(x) > f
PHC
),
lethal optimization
0 1 2 3 4 5 6 7 8 9 10
#Neighbors K
0
0.5
1
1.5
2
Model parameter α
10
-5
10
-4
10
-3
10
-2
10
-1
10
0
10
1
10
2
Ratio P(f(x) > f
TGA
)/P(f(x) > f
TGA,TS=2,p
c
=0.0,p
m
=1/N
),
non-lethal optimization
0 1 2 3 4 5 6 7 8 9 10
#Neighbors K
0
0.5
1
1.5
2
Model parameter α
10
-1
10
0
10
1
10
2
Ratio P(f(x) > f
TGA
)/P(f(x) > f
PHC
),
non-lethal optimization
0 1 2 3 4 5 6 7 8 9 10
#Neighbors K
0
0.5
1
1.5
2
Model parameter α
10
-3
10
-2
10
-1
10
0
10
1
10
2
10
3
Figure 3: (Top) Plots comparing the relative performance of TGA (default parameters) and TGA with TS = 2, p
c
= 0.0, p
m
=
1/N, in a lethal (top left) and non-lethal environment (top right). Darker shades indicate that TGA with TS = 2, p
c
= 0.0, p
m
=
1/N performs better. (Bottom) Plots comparing the relative performance of TGA and PHC (both using default parameters) in a
lethal (bottom left) and non-lethal environment (bottom right). Darker shades indicate that PHC performs better. Performance
is plotted as a function of K and α in all cases. Relative performance is calculated as P( f(x) > f
A
)/P( f (x) > f
B
) where P is a
sample probability estimated from 10000 samples and f (x) is a random draw from the search space, while f
A
and f
B
represent
the best fitness obtained by the two algorithms.
traditional setup on rugged landscapes with structure.
Let us now analyze how the search algorithms
fare with different parameter settings. Table 2 shows
the average best solution fitness obtained by different
algorithm setups in a lethal and non-lethal environ-
ment (values in parenthesis) on NKα landscapes with
K = 4,α = 2.0 (top table) and K = 10,α = 2.0 (bot-
tom table), respectively. Note that due to the nature
of NKα landscapes (particularly due to the absence of
plateaus or neutral networks in the landscapes), there
was only a single best solution in the population at
each generation. For RBS, this has the effect that
the performance is independent of the crossover rate
because crossover is applied to identical solutions.
The two tables largely confirm the observation made
above: while a relatively high degree of population
diversity and randomness in the solution generation
process may be beneficial in a non-lethal environment
(which is particularly true for rugged landscape as can
be seen from the bottom table), it is rather a draw-
back in a lethal environment as it may increase the
risk of generating lethal solutions and subsequently
limit the effectiveness of evolutionary search. More
precisely, for a lethal environment, the tables indicate
that one should use a GA instead of a PHC, and re-
duce randomness in the solution generation process
by avoiding crossover (generally), and using constant
mutation as well as a relatively large tournament size.
The poor performance of Poisson mutation, which is
a typical mutation mode for GAs, is related to situa-
tions where an over-averaged number of solution bits
is flipped at once. Such variation steps lead to off-
spring that are located in the search space far away
from their non-lethal parents, which can in turn in-
crease the probability of generating lethal solutions.
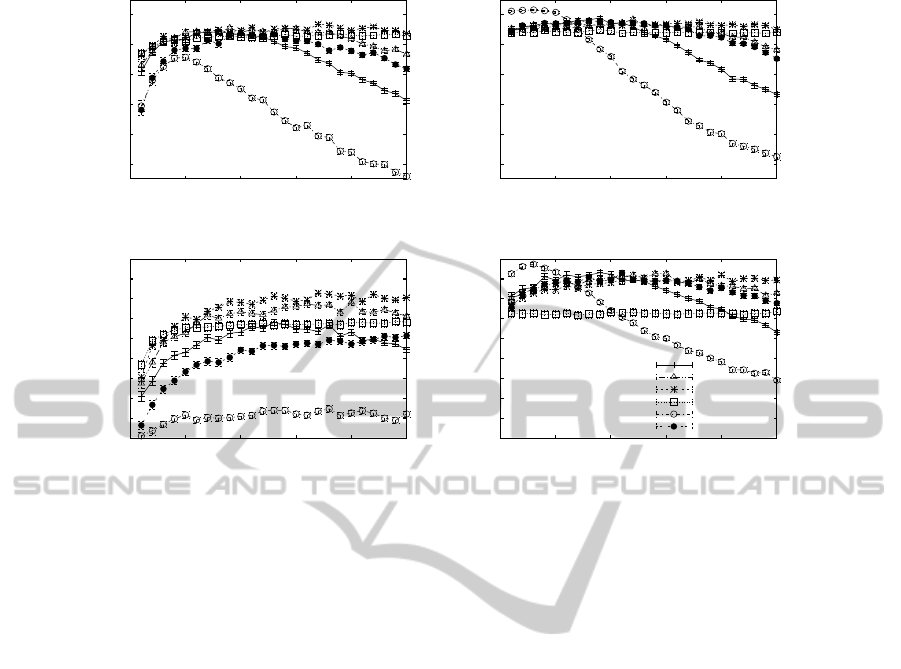
Finally, Figure 4 analyzes whether evolving a
large set of entities for a small number of genera-
tions yields better performance than evolving only a
few entities for many generations; i.e. how is the
trade-off between the initial population size µ
0
and
the maximum number of generations G. For a non-
lethal environment (right plots), we make two obser-
EVOLUTIONARY SEARCH IN LETHAL ENVIRONMENTS
69
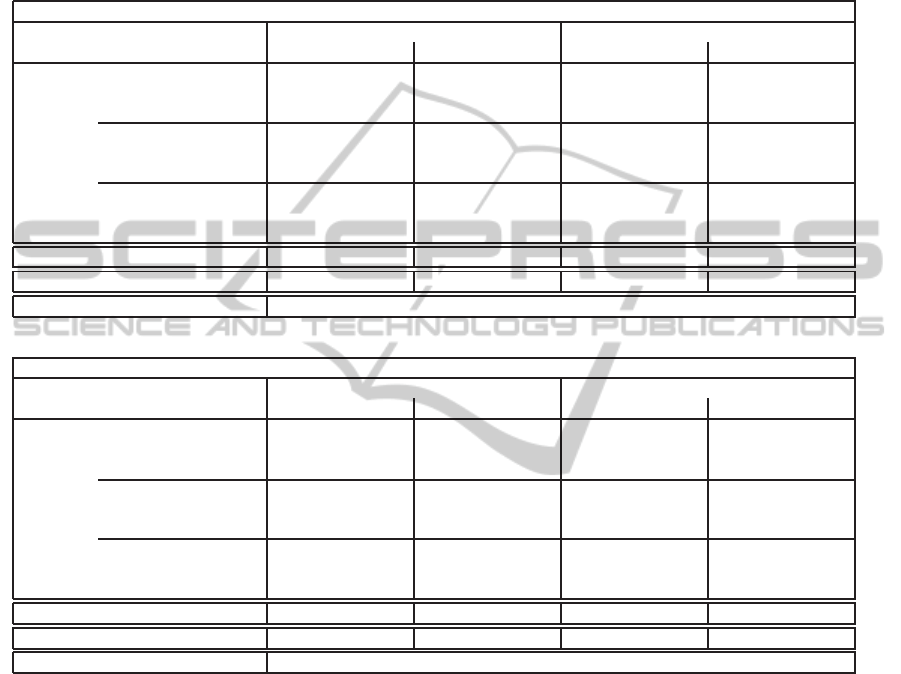
Table 2: The table shows the average best solution fitness found in a lethal environment after G = 200 generations, and
in parenthesis, the fitness found in a non-lethal environment, for different algorithm setups on K = 4,α = 2.0 (top) and
K = 10,α = 2.0 (bottom); the results of random sampling were obtained by generating G × µ
0
= 200× 30 = 6000 solutions
per run at random and averaging over the best solution fitness values found. The numbers in the subscript indicate the rank
of the top five algorithm setups within the respective environment and fitness landscape configuration. We highlighted all
algorithm setups (among the top five) in bold face that are not significantly worse than any other setup. A Friedman test
revealed a significant difference between the search algorithm setups in general, but differences among the individual setups
were tested for in a post-hoc analysis using (paired) Wilcoxon tests (significance level of 5%) with Bonferroni correction.
K = 4, α = 2.0
Constant mutation Poisson mutation
d = 1 d = 2 p
m
= 0.5/N p
m
= 1.0/N
TGA
TS = 1
p
c
= 0.0 0.722
4
(0.7297
4
) 0.6998 (0.7286) 0.7174 (0.7272) 0.7141 (0.7285)
p
c
= 0.25 0.7188 (0.7272) 0.6969 (0.7296
5
) 0.7128 (0.7275) 0.7029 (0.7296)
p
c
= 0.5 0.713 (0.7279) 0.6901 (0.7287) 0.7091 (0.7276) 0.7078 (0.7277)
TS = 2
p
c
= 0.0 0.7223
3
(0.7269) 0.7053 (0.7287) 0.7198 (0.7271) 0.7175 (0.7275)
p
c
= 0.25 0.7217 (0.7263) 0.7039 (0.7284) 0.7172 (0.7263) 0.7119 (0.7275)
p
c
= 0.5 0.7178 (0.7261) 0.7003 (0.728) 0.7176 (0.7268) 0.7135 (0.727)
TS = 4
p
c
= 0.0 0.7218
5
(0.7266) 0.7107 (0.726) 0.7203 (0.7269) 0.7184 (0.7269)
p
c
= 0.25 0.7238
1
(0.7257) 0.7085 (0.7254) 0.7196 (0.7273) 0.7169 (0.7278)
p
c
= 0.5 0.7224
2
(0.7251) 0.7101 (0.7256) 0.7172 (0.7263) 0.7167 (0.726)
RBS 0.7179 (0.7226) 0.7129 (0.723) 0.7177 (0.7229) 0.7172 (0.7223)
PHC 0.7117 (0.7334
2
) 0.6843 (0.7271) 0.7033 (0.7343
1
) 0.6912 (0.7329
3
)
Random sampling 0.6197 (0.6358)
K = 10,α = 2.0
Constant mutation Poisson mutation
d = 1 d = 2 p
m
= 0.5/N p
m
= 1/N
TGA
TS = 1
p
c
= 0.0 0.6918 (0.7317) 0.6637 (0.7383
4
) 0.6812 (0.7338) 0.6767 (0.7356
5
)
p
c
= 0.25 0.6752 (0.7324) 0.6546 (0.7332) 0.6623 (0.7299) 0.6578 (0.7324)
p
c
= 0.5 0.6653 (0.7298) 0.6472 (0.7331) 0.6478 (0.7264) 0.6472 (0.7287)
TS = 2
p
c
= 0.0 0.702
4
(0.7299) 0.6697 (0.7318) 0.6938 (0.7307) 0.6821 (0.7305)
p
c
= 0.25 0.6897 (0.7277) 0.662 (0.732) 0.6717 (0.7281) 0.6643 (0.7302)
p
c
= 0.5 0.6768 (0.7288) 0.6537 (0.7296) 0.6607 (0.7263) 0.659 (0.7274)
TS = 4
p
c
= 0.0 0.7072
2
(0.7275) 0.677 (0.7294) 0.6981
5
(0.7302) 0.6902 (0.7272)
p
c
= 0.25 0.6979 (0.7294) 0.6701 (0.7298) 0.6834 (0.7279) 0.6805 (0.7263)
p
c
= 0.5 0.6936 (0.7265) 0.6635 (0.7296) 0.6715 (0.7258) 0.67 (0.7255)
RBS 0.7102
1
(0.7207) 0.6871 (0.7219) 0.7056
3
(0.7217) 0.6987 (0.7215)
PHC 0.6651 (0.7434
2
) 0.6476 (0.7345) 0.6568 (0.7438
1
) 0.6517 (0.7421
3
)
Random sampling 0.632 (0.6524)
vations: (i) best performance is achieved with PHC
using a rather small population size of about µ
0
= 30,
and (ii) for population sizes µ
0
> 50 one should use a
GA whereby the applied tournament size or selection
pressure should increase with the population size. As
the population size increases, the maximum number
of generations G available for optimization is simply
insufficient for PHC or a GA with low selection pres-
sure and/or much randomness in the solution genera-
tion process to converge quickly to a high-quality part
of the search space. However, on the other hand, a
GA with too much selection pressure may cause the
population to get stuck at local optima, especially on
rugged fitness landscapes (see result of RBS in the
bottom right plot). In a lethal optimization scenario
(left plots), a GA clearly outperforms PHC (regard-
less of the population size) due to the reasons men-
tioned above. Furthermore, unlike to the non-lethal
case, we observe that a GA should use an above-
average large tournament size already for small popu-
lation sizes of µ
0
> 30. Similar to the non-lethal case,
however, there is a saturation point with RBS, beyond
which a further increase in the population size has no
effect (see e.g. bottom left plot).
4 SUMMARY AND
CONCLUSIONS
This paper has conducted an initial investigation of
the impact of lethal solutions (here modeled as candi-
ECTA 2011 - International Conference on Evolutionary Computation Theory and Applications
70
0.67
0.68
0.69
0.7
0.71
0.72
0 50 100 150 200 250
Average best solution fitness
Initial population size µ
0
K=4, α=2.0, lethal optimization
0.67
0.68
0.69
0.7
0.71
0.72
0 50 100 150 200 250
Average best solution fitness
Initial population size µ
0
K=4, α=2.0, non-lethal optimization
0.66
0.67
0.68
0.69
0.7
0.71
0.72
0.73
0.74
0.75
0 50 100 150 200 250
Average best solution fitness
Initial population size µ
0
K=10, α=2.0, lethal optimization
0.66
0.67
0.68
0.69
0.7
0.71
0.72
0.73
0.74
0.75
0 50 100 150 200 250
Average best solution fitness
Initial population size µ
0
K=10, α=2.0, non-lethal optimization
TGA, TS=1
TGA, TS=2
TGA, TS=4
RBS
PHC
TGA, TS=2,p
c
=0.0,p
m
=1/N
Figure 4: Plots showing the average best solution fitness obtained by different search algorithms in a lethal environment (left)
and non-lethal environment (right) on NKα landscapes with K = 4,α = 2.0 (top) and K = 10,α = 2.0 (bottom) as a function
of the initial population size µ
0
; the maximum number of fitness evaluations available for optimization was fixed at 6000
(corresponding to the default setting G = 200,µ
0
= 30), i.e. G = ⌊6000/µ
0
⌋. If not otherwise stated, an algorithm used a
constant mutation mode with d = 1 and no crossover, i.e. p
c
= 0.0.
date solutions below a certain fitness threshold) on
evolutionary search. In essence, the effect of evaluat-
ing a lethal solution is that the solution is immediately
removed from the population and the population size
is reduced by one. This kind of scenario can be found
in Natural evolution, where the presence of lethal mu-
tants may promote robustness over evolvability, but it
is also a characteristic of certain closed-loop evolu-
tion applications, for example, in autonomous robots
or nano-technologies. When faced with this kind of
scenario in optimization, the challenge is to discover
innovative and fit candidate solutions without reduc-
ing the population too rapidly.
Our analysis has been focused on the following
three main aspects: (i) analyzing the impact of lethal
solutions on a standard evolutionary algorithm (EA)
framework, (ii) determining challenging fitness land-
scape topologies in a lethal environment, and (iii) tun-
ing EAs to perform well within a lethal optimiza-
tion scenario. Generally, the presence of lethal so-
lutions can affect that performance of EAs, but the
largest (negative) impact was observed for fitness
landscapes that are rugged and possess some struc-
ture in the sense that some solution bits are more im-
portant than others; in terms of our test functions,
which were NKα landscapes, this type of landscape
corresponds to large values of K and α. For this
fitness landscape topology, we obtained best perfor-
mance in a non-lethal environment using a small pop-
ulation of stochastic hill-climbers. In a lethal environ-
ment, however, significantly better results were ob-
tained using an EA that limits randomness in the so-
lution generation process by employing elitism, a rel-
atively large selection pressure, a constant mutation
mode (i.e. flipping exactly one solution bit as op-
posed to flipping each bit independently with some
low probability), and no or a small crossover rate.
Also, in a lethal optimization scenario, we observed
that an EA that evolves a large set of entities (e.g. au-
tonomous robots or software) for a small number of
generations performs better than one that evolves a
few entities for many generations. The practical im-
plication of this is that, if possible, a larger budget
should be allocated for the production of the entities
in the first place rather than for attempting to prolong
the testing or optimization phase.
5 FUTURE WORK
Our study has of course been very limited, and there
remains much else to learn about optimization prob-
lems subject to lethal solutions and search policies
EVOLUTIONARY SEARCH IN LETHAL ENVIRONMENTS
71

for dealing with these problems. Our current re-
search is looking at the design and tuning of intelli-
gent search policies. We have experimented with an
EA that learns offline, e.g. using reinforcement learn-
ing, when to switch between different selection and
variation operators settings (online) during the opti-
mization (here we used an approach similar to (All-
mendinger and Knowles, 2011)); this EA can yield
better performance than a static or non-learning EA.
It might be worth mentioning that a well-performing
policy learnt offline by this EA is one that increases
randomness in the solution generation process if and
only if the optimization is in the final stages and the
remaining population of reasonable size.
Alternatively to a learning approach, an EA may
be augmented with a strategy that uses assumptions of
local fitness correlation to pre-screen the designs and
forbids the upload of potential lethals. Such a strat-
egy is similar to brood selection with repair and/or
some fitness approximation schemes used in EAs to
filter solutions before evaluation (Walters, 1998; Jin,
2005).
Finally, analyzing the effect of lethal solutions and
search policies on different and perhaps more realis-
tic fitness landscapes than NKα landscapes, e.g. ones
with neutral plateaux, is another avenue we are pur-
suing. In the further future, we might enjoy trying
out our strategies on real lethal environments in au-
tonomous robots, nano-technologies or similar.
REFERENCES
Allmendinger, R. and Knowles, J. (2011). Policy learning
in resource-constrained optimization. In Proceedings
of GECCO, pages 1971–1978.
Barnett, L. (2001). Netcrawling-optimal evolutionary
search with neutral networks. In Proceedings of CEC,
pages 30–37.
Bullock, S. (2003). Will selection for mutational robustness
significantly retard evolutionary innovation on neutral
networks? In Artificial Life VIII, pages 192–201.
Calzolari, D., Bruschi, S., Coquin, L., Schofield, J., Feala,
J. D., Reed, J. C., McCulloch, A. D., and Paternostro,
G. (2008). Search algorithms as a framework for the
optimization of drug combinations. PLoS Computa-
tional Biology, 4:1.
Ciuti, G., Donlin, R., Valdastri, P., Arezzo, A., Menciassi,
A., Morino, M., and Dario, P. (2010). Robotic versus
manual control in magnetic steering of an endoscopic
capsule. Endoscopy, 42(2):148–52.
Forrest, S. and Mitchell, M. (1993). Relative building-block
fitness and the building-block hypothesis. In Founda-
tions of Genetic Algorithms 2, pages 109–126.
Harvey, I., Husbands, P., Cliff, D., Thompson, A., and
Jakobi, N. (1996). Evolutionary robotics: The Sussex
approach. Robotics and Autonomous Systems, 20(2-
4):205–224.
Hebbron, T., Bullock, S., and Cliff, D. (2008). NKα:
Non-uniform epistatic interations in an extended NK
model. In Artificial Life XI, pages 234–241.
Hughes, B. D. (1995). Random walks and random environ-
ments. Clarendon Press.
Jin, Y. (2005). A comprehensive survey of fitness approxi-
mation in evolutionary computation. Soft Computing,
9(1):3–12.
Kauffman, S. (1989). Adaptation on rugged fitness land-
scapes. In Lecture Notes in the Sciences of Complex-
ity, pages 527–618.
Klockgether, J. and Schwefel, H.-P. (1970). Two-phase noz-
zle and hollow core jet experiments. In Engineering
Aspects of Magnetohydrodynamics, pages 141–148.
Moglia, A., Menciassi, A., Schurr, M. O., and Dario, P.
(2007). Wireless capsule endoscopy: from diagnostic
devices to multipurpose robotic systems. Biomedical
Microdevices, 9(2):235–243.
Rechenberg, I. (2000). Case studies in evolutionary exper-
imentation and computation. Computer Methods in
Applied Mechanics and Engineering, 2-4(186):125–
140.
Schonfeld, J. (2007). A study of mutational robustness as
the product of evolutionary computation. In Proceed-
ings of GECCO, pages 1404–1411.
Shir, O. M., Emmerich, M., B¨ack, T., and Vrakking, M.
J. J. (2007). The application of evolutionary multi-
criteria optimization to dynamic molecular alignment.
In Proceedings of CEC, pages 4108–4115.
Small, B. G., McColl, B. W., Allmendinger, R., Pahle,
J., L´opez-Castej´on, G., Rothwell, N. J., Knowles, J.,
Mendes, P., Brough, D., and Kell, D. B. (2011). Ef-
ficient discovery of anti-inflammatory small molecule
combinations using evolutionary computing. Nature
Chemical Biology, (to appear).
Syswerda, G. (1989). Uniform crossover in genetic algo-
rithms. In Proceedings of ICGA, pages 2–9.
Thompson, A. (1996). Hardware Evolution: Automatic de-
sign of electronic circuits in reconfigurable hardware
by artificial evolution. PhD thesis, University of Sus-
sex.
Walters, T. (1998). Repair and brood selection in the travel-
ing salesman problem. In Proceedings of PPSN, pages
813–822.
Watson, R. A., Ficici, S. G., and Pollack, J. B. (2002). Em-
bodied evolution: Distributing an evolutionary algo-
rithm in a population of robots. Robotics and Au-
tonomous Systems, 39(1):1–18.
Zykov, V., Mytilinaios, E., Adams, B., and Lipson,
H. (2005). Self-reproducing machines. Nature,
435(7039):163–164.
ECTA 2011 - International Conference on Evolutionary Computation Theory and Applications
72
