
DEVELOPMENTAL PLASTICITY IN CARTESIAN GENETIC
PROGRAMMING BASED NEURAL NETWORKS
Maryam Mahsal Khan, Gul Muhammad Khan
Electrical Engineering Department, University of Engineering and Technology, Peshawar, Pakistan
Julian F. Miller
Department of Electronics, University of York, York, U.K.
Keywords:
Generative and developmental approaches, NeuroEvolutionary algorithms, Pole balancing.
Abstract:
This work presents a method for exploiting developmental plasticity in Artificial Neural Networks using Carte-
sian Genetic Programming. This is inspired by developmental plasticity that exists in the biological brain
allowing it to adapt to a changing environment. The network architecture used is that of a static Cartesian
Genetic Programming ANN, which has recently been introduced. The network is plastic in terms of its dy-
namic architecture, connectivity, weights and functionality that can change in response to the environmental
signals. The dynamic capabilities of the algorithm are tested on a standard benchmark linear/non-linear control
problems (i.e. pole-balancing).
1 INTRODUCTION
Natural neural systems are not static in nature. They
interact and respond to environmental factors. Such
dynamic interactions lead to modification or devel-
opment of the system with time. A static system
can be trained to give a specific response to a set
of environmental stimuli however it cannot learn or
adapt to a changing environment. To do this one
requires a network that develops at runtime. Evo-
lutionary computation based algorithms have proved
to be effective at finding a particular solution to a
computational problem, however, if the problem do-
main is slightly changed the evolved network is un-
able to solve the problem. Catastrophic forgetting
(CF) is the phenomenon in which a trained artificial
neural network loses its accuracy when the network
is trained again on a different computational prob-
lem (McCloskey and Cohen, 1989; Ratcliff, 1990;
Sharkey and Sharkey, 1995; Ans et al., 2002; French,
1994). Forgetting is inevitable if the learning resides
purely in the weights of fixed connections.
Our aim is to find a set of computational func-
tions that encode neural network architecture with an
ability to adapt to the changing environment as a re-
sult of development. Such plastic neural networks are
different from conventional ANN models as they are
constantly adjusting both topology and weights in re-
sponse to external environmentalsignals. This means,
that they can grow new networks of connections when
the problem domain requires it. The main motiva-
tion for such a new model is to arrive at a plastic
neural network model that can produce a single neu-
ral network that can solve multiple linear/non-linear
problems (i.e. not suffer from a phenomenon called
catastrophic interference). We think that this can be
acheived by arriving at a neural network model in
which new neural sub-structures automatically come
into existence when the network is exposed to new
learning scenarios.
In this paper we introduce a plastic neural network
based on the representation of Cartesian Genetic Pro-
gramming (Miller and Thomson, 2000) (PCGPANN).
The performance of this algorithms is tested on the
standard benchmark problem of pole-balancing for
both linear and non-linear conditions.
In Section 2 we will briefly review the current de-
velopmental algorithms presented to date. Section 3
gives an overview of Cartesian Genetic Programming
(CGP). In section 4, we will provide an overview
of the standard CGPANN algorithm. Section 5 de-
scribes how self-modification or developmental plas-
ticity could be embedded in the standard CGPANN
algorithm. Section 6 - 8 is the application of the algo-
449
Mahsal Khan M., Muhammad Khan G. and F. Miller J..
DEVELOPMENTAL PLASTICITY IN CARTESIAN GENETIC PROGRAMMING BASED NEURAL NETWORKS.
DOI: 10.5220/0003615204490458
In Proceedings of the 8th International Conference on Informatics in Control, Automation and Robotics (ANNIIP-2011), pages 449-458
ISBN: 978-989-8425-74-4
Copyright
c
2011 SCITEPRESS (Science and Technology Publications, Lda.)

rithm on a standard benchmark problem - single and
double pole-balancing along with results and discus-
sion. Section 9 concludes with the findings and future
work.
2 RELATED WORK
GP has been used in a number of ways to produce
ANNs. The simplest of all is using direct encoding
schemes in which GP is used to encode the weights
of ANN or architecture or both. In direct encoding
schemes each and every unit in phenotype is specified
in the genotype. Specifying each connection of ANN
in genotype is commonly referred to as ‘structural en-
coding’ (Hussain and Browse, 2000).
Another important form of encoding that is used
to develop ANNs is Grammar encoding. There are
two types of grammar encoding schemes: develop-
mental grammatical encoding and derivation gram-
matical encoding. Kitano used developmental gram-
matical encoding scheme for development of ANN
(Kitano, 1990). In this, genes describe the grammat-
ical rule that is used for development of the ANN. In
derivation grammatical encoding, the gene contains
a derivation sequence that specify the whole network
(Gruau, 1994; Jacob and Rehder, 1993).
Cellular Encoding (CE) devised by Gruau et.al
(Gruau et al., 1996) is a developmental neuroevolu-
tionary algorithm. Weights, connection, graph re-
writing rules are evolved based on the evolutionary
algorithm. CE transforms graph in a manner that con-
trols cell division which grows into an ANN. CE was
applied on a control problem of balancing single and
double poles. This technique was found to scale bet-
ter and was effective at optimizing both architecture
and weights together.
A model proposed by Nolfi et.al maps a genotype
into a phenotype during the existence of the individ-
ual. The life expectancy of the individual is affected
by the external environment and the genotype itself
(Nolfi et al., 1994). The two dimensional neural net-
work adapts during their lifetime as environmental
conditions affect the growth of the axons. The neu-
rons had only upward growing axons with no den-
drites.
The development of a neural architecture from
a single cell was proposed by Cangelosi. In this
model the cell undergoes cell division and migra-
tion to produce a two dimensional neural architecture
(Cangelosi et al., 1994). The rules for both the pro-
cess of cell division and migration is specified in the
genotype; see (Dalaert and Beer, 1994; Gruau, 1994)
for related approaches.
Another developmentalmodel that evolves param-
eters based on genetic algorithm was proposed by
Rust and Adams. The evolved parameters represented
neurons that were biologically realistic. They also in-
vestigated the influence of dependent mechanisms on
growing morphologies of neuron (Rust et al., 2000).
Karl Sims used a graph based GP approach to
evolve virtual robotic creatures. The neural architec-
ture for controlling the body parts of the robots were
genetically determined (Sims, 1994).
Rogen et.al proposed a developmental spiking
ANN cellular model. Each cell is composed of an ex-
citory or inhibitory weight input neurons. The neuron
triggers once a certain threshold is reached and goes
into a short refractory period. Because of the leak-
age mechanism in spiking ANN, the threshold of the
membrane reduces with time (Roggen et al., 2007).
Rivero, evolved a specially designed tree-
representation which could produced an ANN graph
(Rivero et al., 2007). He tested his network on a num-
ber of data mining applications.
Harding et al. adapted the graph-based genetic
programming approach, known as Cartesian Genetic
Programming (CGP) by introducing a number of in-
novations. Foremost of which was the introduction
of self-modifying functions. The method is called
self-modifying CGP (SMCGP) (Harding et al., 2010).
Self-modifying functions allow an arbitrary number
of phenotypes to be encoded in a single genotype.
SMCGP was evaluated on a range of problems e.g.
digital circuits, pattern and sequence generation and,
notably, was found to produce general solutions to
some classes of problem (Harding et al., 2010).
Khan et al. presented a neuro-inspired devel-
opmental model using indirect encoding scheme to
develop learning neural architectures (Khan et al.,
2007). They used cartesian genetic programming to
encode a seven chromosome neuron genotype. The
programs encoded controlled the processing of sig-
nals and development of neural structures with mor-
phology similar in certain ways to biological neurons.
The approach has been tested on a range of AI prob-
lems including: Wumpus world, Solving Mazes and
the Game of Checkers (Khan et al., 2007).
The N-gram GP system is extended to incorporate
developmental plasticity known as ‘Incremental Fit-
ness based Development (IFD)’ proposed by McPhee
et.al (Nicholas F.McPhee and Poli, 2009). Both N-
gram and IFD have been applied on various regres-
sion problems where IFD has been found producing
better results.
HyperNEAT is one of the indirect encoding strate-
gies of developmental ANNs (Stanley et al., 2009).
HyperNEAT uses generative encoding and the basic
ICINCO 2011 - 8th International Conference on Informatics in Control, Automation and Robotics
450
evolutionary principles based on the NEAT (Neuro
Evolution of Augmented Topology) algorithm (Stan-
ley and Miikkulainen, 2002). In HyperNEAT the
weights(inputs) are presented to an evolved program
called a Compositional Pattern Producing Networks
(CPPNs). The CPPN takes coordinates of neurons in
a grid and outputs the corresponding weight for that
connection. The algorithm has been applied to Robot
locomotion. Regular quadruped gaits in the legged lo-
comotion problem have been successfully generated
using this algorithm (Clune et al., 2008).
Synaptic plasticity in Artificial neural network can
be introduced by using local learning rules that mod-
ify the weights of the network at runtime (Baxter,
1992). Floreano and Urzelai have demonstrated that
evolving network with synaptic plasticity can solve
complex problemsbetter than recurrent networks with
fixed-weights (Floreano and Urzelai, 2000). A num-
ber of researchers have investigated and compared the
performance of plastic and recurrent networks obtain-
ing mixed results with either of them performing bet-
ter in various problem domains (for a review see (Risi
et al., 2010)). One of the interesting features of the ap-
proach we propose is that, unlike previous approaches
investigated, it has both synaptic and developmental
plasticity.
3 CARTESIAN GENETIC
PROGRAMMING
Cartesian Genetic Programming (CGP) is an evo-
lutionary technique for evolving graphs (Miller and
Thomson, 2000). The graphs are arranged in the
form of rectangular grids like multi-layer percep-
trons. The graph topology is defined using three pa-
rameters: number of columns, number of rows and
levels-back. The number of nodes in the graph en-
coded in the genotype is the product of the number
of columns and the number of rows. Usually feedfor-
ward graphs are evolved in which inputs to a node are
the primary inputs or outputs from the previous nodes.
The levels-back parameter constrains connections be-
tween nodes. For instance, if levels-back is one, all
nodes can only be connected to nodes immediately
preceeding them, while if levels-back is equal to the
number of columns of nodes (primary inputs are al-
lowed to disobey this constraint) the encoded graphs
could be an arbitrary feed-forward graph. Genotypes
consist of a fixed length string of integers (referred to
as genes). These genes represents nodes with inputs
and functions. Functions can be any linear/non linear
functions. The output of the genotype can be from
any node or the primary inputs. CGP generally uses a
1+4 evolutionary strategy to evolve the genotype. In
this the parent genotype is unchanged and 4 offspring
are produced by mutating the parent genotype. It is
important to note that when genotypes are decoded
nodes may be ignored as they are not connected in
the path from inputs to outputs. The genes associated
with such nodes are non-coding. This means that the
active final graph encoded could consist of a pheno-
type containing any subset of number of nodes de-
fined in the genotype. Mutating a genotype can thus
change the phenotype either not at all or drastically.
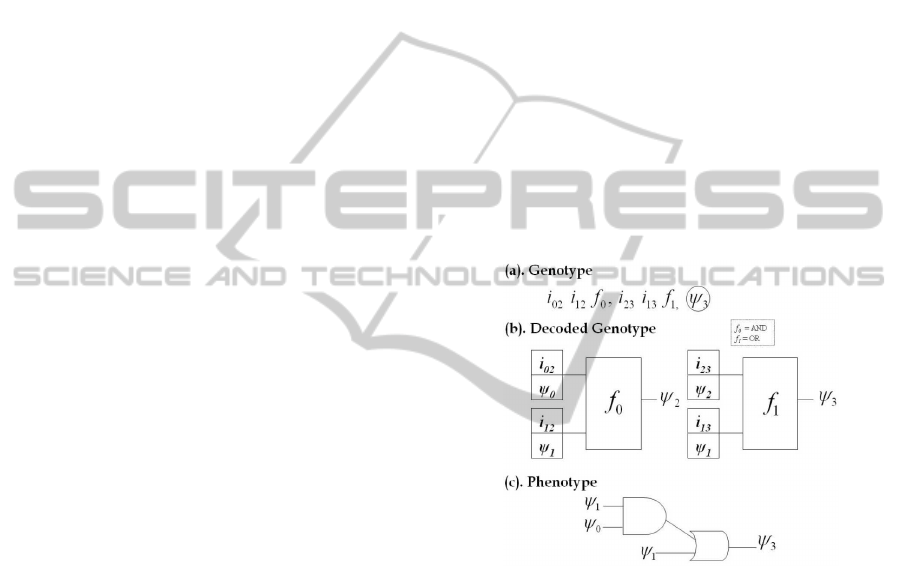
We give a simple example of CGP in Figure.1
where 1(a) shows a 1x2 (rows x columns) geno-
type for a problem with 2 binary inputs ‘ψ
0,1
’ and
1 binary output. The inputs to each node is repre-
sented as i
mn
where ‘m’ represents the input from
and ‘n’ is the input to the node. The functions ‘f
n
’
used are logical ‘OR’ and logical ‘AND’. Figure.1(b)
shows the graphical representation of the genotype
in Figure.1(a). The resultant phenotype shown in
Figure 1: (a) CGP based Genotype for a problem with 2
binary inputs ψ
0
and ψ
1
. Single program output is taken
from the last gene (circled). (b) Decoding of the genotype.
(c) Phenotype represented in terms of a digital circuit.
Figure.1(c) can be represented as a logical equation
in Eq.(1) where ‘+’ represent OR and ‘.’ represents
AND operation.
ψ
3
= ψ
1
.ψ
0
+ ψ
1
(1)
4 CGP EVOLVED ANN
The representation of CGP has been exploited in gen-
erating static neural architectures. Both feedforward
and recurrent (FCGPANN , RCGPANN) based net-
works have been designed and tested on various ap-
plications (Khan et al., 2010b; Khan et al., 2010a).
DEVELOPMENTAL PLASTICITY IN CARTESIAN GENETIC PROGRAMMING BASED NEURAL NETWORKS
451
In CGPANN, along with standard CGP genes (in-
puts, functions) weight and switch genes are also in-
troduced. The weight values are randomly assigned to
each input with values from (−1,+1). Switch genes
are either ‘1’ or ‘0’ where ‘1’ is a connected and ‘0’ a
disconnected input. The genotype is then evolved to
obtain best topology, optimal weights and combina-
tion of functions.
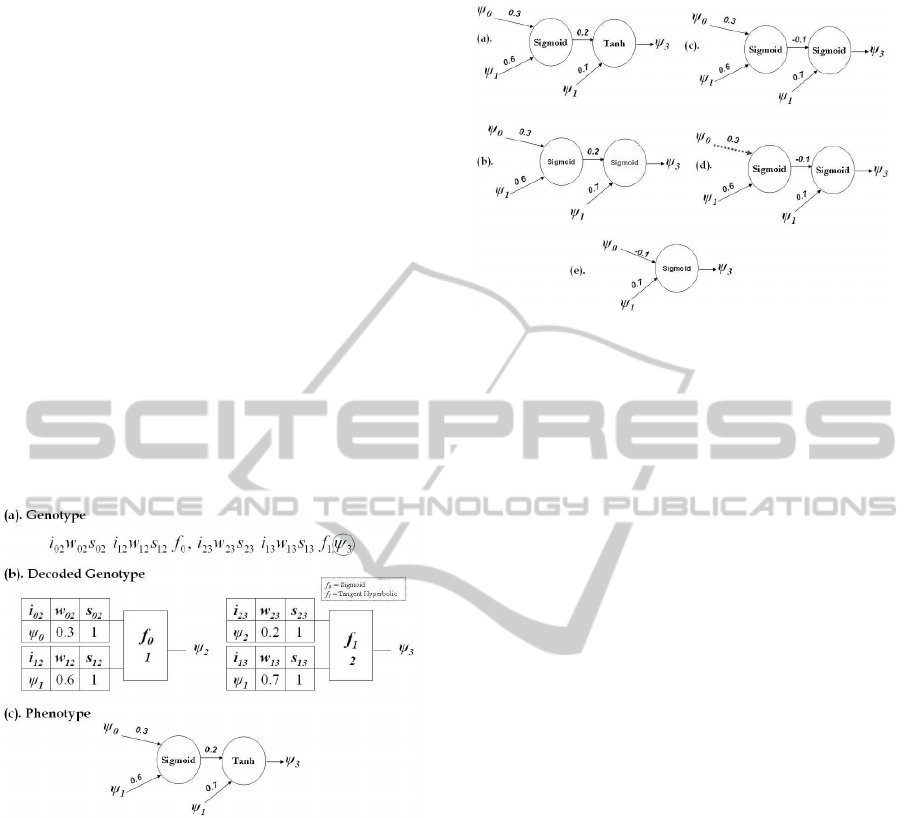
Figure.2 is an example of a CGPANN genotype
with two primary inputs ψ
0,1
and one output. The
activation function f
n
used are sigmoid and tangent
hyperbolic. The inputs to each node is represented as
i
mn
. Similarly weights w
mn
are assigned to each in-
put. Whether an input is connected or not is encoded
as a switching gene s
mn
. Figure.2(a) is the genotype
of a 1x2 architecture with the output taken from node
ψ
3
. Figure.2(b) is the graphical view of the genotype
with values assigned e.g. the weight assigned to the
input i
12
supplied to node 2 is w
12
= 0.6. The pheno-
type of the genotype is shown in Figure.2(c) which is
mathematically expressed in Eq.(2).
Figure 2: (a) Genotype of a 1x2 architecutre with two in-
puts. (b) Decoded Genotype. (c) Phenotype represented as
ANN of the genotype.
ψ
3
= Tanh(0.7ψ
1
+ 0.2sig(0.3ψ
0
+ 0.6ψ
1
)) (2)
5 PLASTIC CGP NEURAL
NETWORK
A plastic CGPANN (PCGPANN) is a developmental
form of Cartesian Genetic Programming ANN algo-
rithm. The basic structure and representation of the
PCGPANN is similar to the CGPANN structure as
shown in Figure. 2. In the PCGPANN method, an
additional output gene is added to the genotype. It is
used for making developmental decisions. According
to whether the output value is less than zero or above,
Figure 3: PCGPANN - Plastic Development of Neural Net-
work Parameters (a) Initial Phenotype. (b) Mutation of
Function. (c) Mutation of Weight. (d) Mutation of Switch.
(e) Mutation of Input.
a ‘mutation’ of the genotype is introduced to create
a new phenotype. In this way, a single genotype can
evoke an unlimited series of phenotypes depending
on the real-time output of the program encoded in the
genotype. The modification of the genotype results
in different phenotypic structures. The mutation pro-
duced is randomly chosen from the types listed below.
It should be noted that some gene changes may result
in no phenotypic change as in CGP many genes are
redundant. The phenotypic mutation of neural net-
work can be any of the following: inputs, weights,
switches,outputs and activation functions. Genes are
mutated and assigned a valid value based on sets of
constraints.
• If a gene represents a function then it is replaced
by any random function selected from 1 to total
number of functions (n
f
) available to the system.
• If the gene represents a weight is replaced by a
weight value between (-1, +1).
• If the gene is a switching gene then it is comple-
mented.
• Node input genes are mutated by assigning an in-
put value such that the levels-back parameter is
still obeyed.
• An output can be assigned any random value from
1 to total number of nodes plus inputs presented to
the system.
Figure.3 shows a step by step process of mutation
on a phenotype. Figure.3(a) is the initial phenotype
of the genotype given in Figure.2(c). As in the de-
scribed example the activation functions used were
two - sigmoid and tangent hyperbolic. Thus the geno-
type has only two functions to randomly choose from.
ICINCO 2011 - 8th International Conference on Informatics in Control, Automation and Robotics
452

In Figure.3(b) the function is mutated from tangent
hyperbolic to sigmoid . The mathematical expression
of the updated genotype is thus changed as shown in
Eq.(3).
ψ
3
= sig(0.7ψ
1
+ 0.2sig(0.3ψ
0
+ 0.6ψ
1
)) (3)
Figure.3(c) represents a mutated phenotype where the
value of weight is changed from 0.2 to -0.1 expressed
in Eq.(4).
ψ
3
= sig(0.7ψ
1
− 0.1sig(0.3ψ
0
+ 0.6ψ
1
)) (4)
Similarly Figure.3(d) displays the mutated phenotype
where the switching gene is toggled and switched off
(shown as dotted line). Thus the input is no longer
connected to the node. Mathematically expressed by
Eq.(5).
ψ
3
= sig(0.7ψ
1
− 0.1sig(0.6ψ
1
)) (5)
Mutation of inputs is one of the major tunning pa-
rameter during the developmental process of ANN.
From the phenotype displayed in Figure.3(d), the in-
put to the second node is ψ
1
and an output from the
first node. After mutating the input, the input to the
second node is modified to two primary inputs of the
system ψ
0,1
. The resultant phenotype is displayed
in Figure.3(e) with its function in Eq.(6). Thus dur-
ing runtime different functional equations can be pro-
duced generating desired response.
ψ
3
= sig(−0.1ψ
0
+ 0.7ψ
1
) (6)
Similarly output of any genotype can also be mutated
and changed to another node. In this way inactive
genes present in the genotype can be activated or vice
versa. This in turn effects the phenotypic structure of
the genotype.
It is worth mentioning here that mutation of geno-
type at runtime affects only a single gene where as
mutation from generation to generation is dependent
on the mutation rate fixed at the start of the program,
thus changing the starting genotype for each develop-
ment.
6 CASE STUDY: BALANCING
POLE(S)
Pole balancing also known as inverted pendulum is
a standard benchmark problem in the field of control
theory. The performance of various neuroevolution-
ary algorithms is tested on this problem. Single pole
balancing task consists of a pole attached by a hinged
to a wheel cart while double pole balancing consti-
tutes two poles. In both the cases the track of the cart
Figure 4: (a) Single Pole Balancing. (b) Double Pole Bal-
ancing.
is limited to −2.4 < x < +2.4 as shown in Figure.4.
The objective is to apply force ‘F’ to the cart such
that the cart doesn’t leave the track and the angle of
the pole doesn’t exceed (-12
◦
< θ
1
< +12
◦
) for single
pole and (-36
◦
< θ
1,2
< +36
◦
) for the double pole bal-
ancing task. The controller has to balance the pole(s)
for approximately 30 minutes which corresponds to
100,000 time steps. Thus the neural network must ap-
ply force to the cart to keep the pole(s) balanced for as
long as possible. The input to the system are the angle
of pole(s) (θ
1,2
), angular velocity of pole (
˙
θ
1,2
), posi-
tion of cart (x) and velocity of cart (˙x). Equations that
are used to compute the effective mass of poles, accel-
eration of poles, acceleration of cart, effective force
on each pole, the next state of angle of poles, veloc-
ity of poles, position of cart and velocity of cart along
with the parameters defined for both control problems
can be found in (Khan et al., 2010a).
7 EXPERIMENTAL SETUP
PCGPANN genotypes are generated for various net-
work sizes for both single and double pole balanc-
ing task. A mutation rate of µ
r
= 10% is used for
all the networks. The activation functions f
n
used
are sigmoid and tangent hyperbolic. Each genotype
is initialized with random weights ‘w’ from -1 to +1,
switching ‘s’ values of 0 or 1 and random inputs ‘ψ
i
’.
The number of outputs ‘O
i
’ from the system are 2 i.e.
O
1
and O
2
. Both the outputs are normalized between
-1 to +1. Output O
1
represents the ‘Force (F)’ that
controls the force applied to the cart and the output
O
2
is associated with the developmental plasticity of
the network. Using one of the outputs of the genotype
for making developmental decision acts as a feedback
to the system for generating plastic changes.
We have simulated the network for two develop-
mental strategies Dev
1
and Dev
2
as shown in Eq.(7)-
(8). Both the strategies are tested on the pole bal-
ancing task for single and double pole(s) with zero
DEVELOPMENTAL PLASTICITY IN CARTESIAN GENETIC PROGRAMMING BASED NEURAL NETWORKS
453
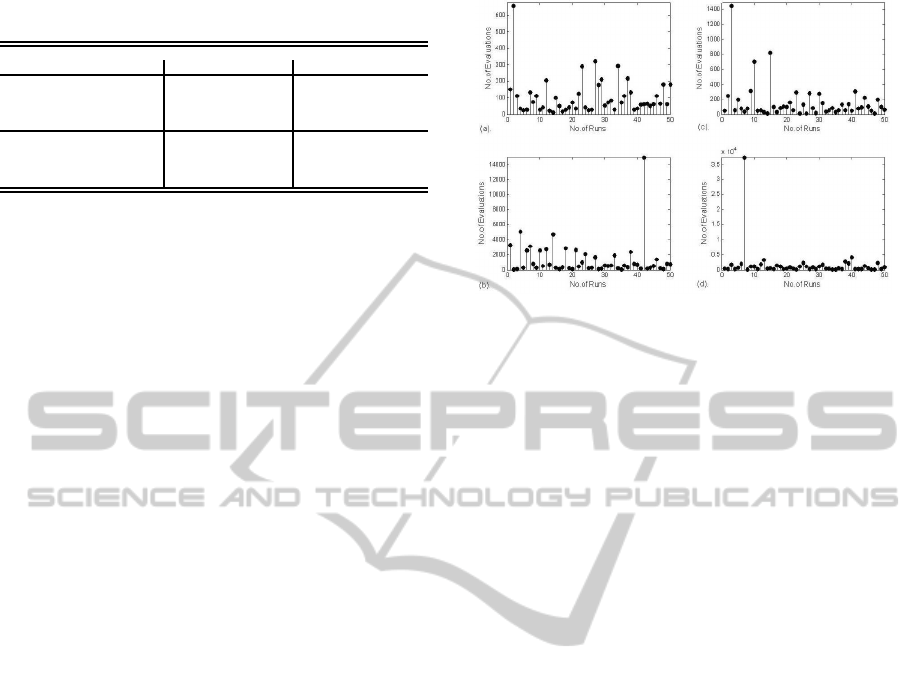
Table 1: Performance of PCGPANN on Single and Double
Poles.
Network Representation SinglePole Evaluations DoublePole Evaluations
Initial State Network Arch. Dev
1
Dev
2
Dev
1
Dev
2
Zero 4x4 221 659 1716 6310
5x5 186 570 1169 5494
10x10 104 519 1337 8315
15x15 168 643 1934 7334
Random 4x4 390 608 1738 3310
5x5 303 387 1981 4033
10x10 157 243 1612 3308
15x15 160 260 1916 4362
and random initial states. The intention is to identify
faster and robust strategies for the problem investi-
gated.
Dev
1
=
(
No −Change -1 ≤ O
2
≤ 0
Mutation− of − Parameters 0< O
2
≤ 1
(7)
Dev
2
=
(
Mutation− of − Parameters -1 ≤ O
2
≤ 0
No −Change 0< O
2
≤ 1
(8)
7.1 Single Pole: Zero and Random
Initial States
In single pole balancing the input to the networks are
position of cart x, velocity of cart ˙x, angle of pole θ
1
and velocity of pole
˙
θ
1
. The number of inputs to each
node is taken as 4, which is equivalent to the number
of inputs to the system.
7.2 Double Pole: Zero and Random
Initial States
Double pole balancing involves balancing an addi-
tional pole. The number of input to the networks are
position of cart x, velocity of cart ˙x, angle of poles θ
1,2
and angular velocity of poles
˙
θ
1,2
. The number of in-
put to each node is taken as 6, which is equivalent to
the number of inputs to the system.
In both the cases (single and double poles), the
networks are generated with initial input values of
zero or with random values of -0.25rad <θ
1
<0.25rad
and -2.4 <x <+2.4 for single pole and -0.6rad <θ
1,2
<0.6rad and -2.4 <x <+2.4 for double poles. The
performance of the PCGPANN algorithm is based on
the average number of balancing attempts (evalua-
tions) for fifty independent evolutionary runs.
8 RESULTS AND DISCUSSIONS
Table 1 represents average number of balancing at-
tempts for inputs starting from initial states of zero
and random states of network with varying network
Figure 5: Variation in the number of evaluations of the
best evolved network’s genotypes for single(a,c) and dou-
ble poles(b,d) with zero and random initial states.
sizes for single and double pole(s) balancing tasks
under both the developmental strategies. The results
are the average of 50 independent evolutionary runs.
Figure.5 shows the variation in the number of evalua-
tions of the best evolved network’s genotypes for sin-
gle/double pole(s) with zero and random initial states.
From the figure it is evident that out of fifty genotypes
only one of them took longer than expected number
of evaluations thus increasing the overall average. It
is observed in all the four cases of Figure.5 that learn-
ing speed is much faster in 98% of the evolutionary
runs. It is observed in Table 1 that the developmen-
tal strategy Dev
1
has performed better than the Dev
2
strategy for both the single and double pole balancing
tasks. From Table 1, the minimum average evalua-
tions of 50 independent runs for the single pole and
double pole was 104 and 1169 respectively. Random
initial states took longer time to develop into a stable
structure as compared to zero initial states for obvi-
ous reasons. For both single and double pole balanc-
ing task, fast learning in terms of minimum average
evaluations has been found in the 10x10 network ar-
chitecture.
Figure.6 represents the pole angles and position of
cart simulated for 30 minutes respectively (100,000
steps are down-sampled to produce 100 steps for
demonstration purpose only) for the single pole and
double pole balancing task. In Figure.6(a) the cart
is moving back and forth to make the pole bal-
anced while minute changes in the angle of pole
is observed. Similarly in case of Figure.6(b) ran-
dom behaviour of movement of both the poles and
the cart is observed for the double pole balancing
task. Pole balancing task is used as a benchmark
by researchers to investigate the performance of al-
gorithms including: Conventional Neuro Evolution
(CNE) (Wieland, 1991), Symbiotic, Adaptive Neural
ICINCO 2011 - 8th International Conference on Informatics in Control, Automation and Robotics
454
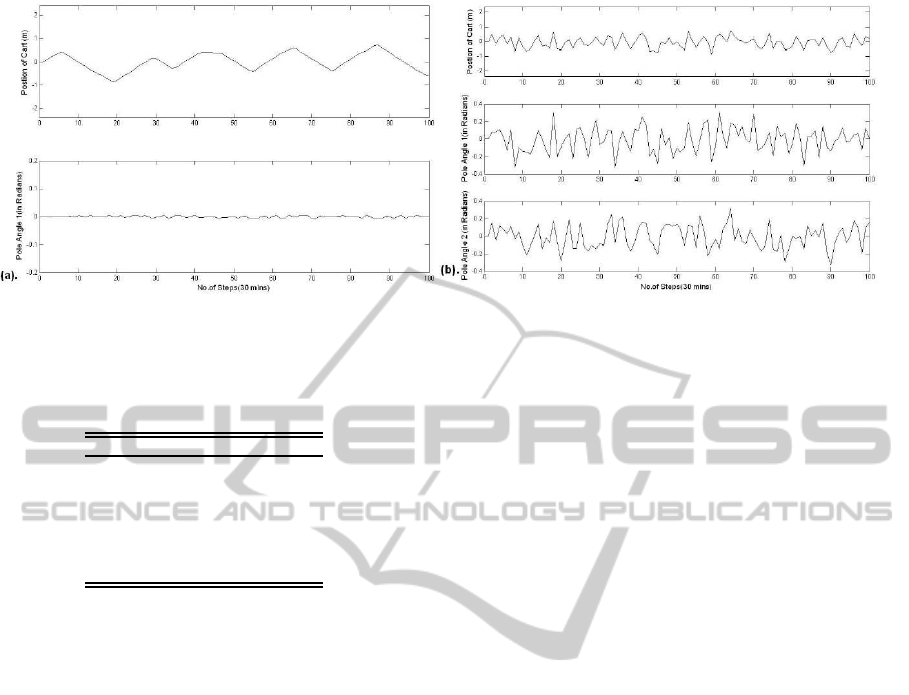
Figure 6: Pole Angle(s) and Position of Cart simulated for 100,000 steps for (a) Single Pole Task (b) Double Pole Task.
Table 2: Comparison of PCGPANN with other neuroevolu-
tionary algorithms Applied on Single and Double Pole Bal-
ancing Task: average number of network evaluations.
Method SinglePole DoublePole
CNE 352 22100
SANE 302 12600
ESP 289 3800
NEAT 743 3600
CoSyNE 98 954
FCGPANN 21 77
PCGPANN 104 1169
Evolution (SANE)(Moriarty, 1997), Enforced Sub-
Population (ESP) (Gomez and Miikkulainen, 1999),
Neuro Evolution of Augmenting Topologies (NEAT)
(Stanley and Miikkulainen, 2002) and Cooperative
synapse neuroevolution (CoSyNE) (Gomez et al.,
2008). All these neuroevolutionary algorithms pro-
duces static ANNs detail of which can be found in
(Khan et al., 2010a). Table 2 represents the compar-
ison of performance of the Plastic CGPANN with all
these neuroevolutionary algorithms for both the sin-
gle and double pole balancing tasks. Developmen-
tal CGPANN has outperformed most of these neu-
roevolutionary algorithms by quite a large margin. As
pointed out by Risi et al. learning a specific task is
easy but learning the strategy to learn and adapting
its architecture to the changing task takes time (Risi
et al., 2010). This is the reason that PCGPANN took
more evaluations to arrive at a solution as compared to
its static counterpart - FeedForward CGPANN (FCG-
PANN)(Khan et al., 2010a).
8.1 Developmental Plasticity
It is observed in the CGP literature that only upto 5%
nodes are usually active at runtime, thus having 95%
of garbage space (inactive nodes) that may be acti-
vated during evolution (Miller and Smith, 2006). As
the overall architecture of the system in PCGPANN
is fixed (5x5, 10x10, etc networks), only the inter-
connectivity of the neurons changes at runtime thus
changing the active morphology of the ultimate phe-
notype. This phenomenon is similar to what is ob-
served in biological brain with different parts of ner-
vous system responding to different sets of input at
runtime (Kandel et al., 2002). In Plastic CGPANN
algorithm neurons are presented in terms of network
sizes where a 5x5 network corresponds to 25 neu-
rons. The target is then to use these neurons like re-
sources in a system in an effective and efficient man-
ner. The structure produced connects and disconnects
during the developmental process generating differ-
ent sets of equations at run time. This corresponds to
the creation of different phenotypes during a genera-
tion, similar to the work done by Nolfi (Nolfi et al.,
1994). Such plasticity embeds efficient utilization of
resources and reduces hardware cost. Till date no
developmental algorithm other than PCGPANN have
been proposed that has excessive neuron structures
that activate and deactivate with time rather fix ar-
chitectures are mostly proposed. Figure.7 shows the
logarithmic graphs for developmental outputs of six
different evolved genotypes for both the developmen-
tal strategies while balancing double poles for 30mins
(100,000 time steps). It is evident from these graphs
that after some early development the ultimate output
of the network causes the network to stay same (no
change decision). It can be seen in Figure.7(a,c,e)that
genotypes produces developmental output with oscil-
lating behaviour at the beginning resulting in change
of phenotypic structure and weights more often. Sim-
ilar behaviour is also observed in biological brain
where most of the learning and development happens
in the immature brain as compared to the adult brain.
Hence changing or mutating parameters at the early
stage has greater probability of convergence to the so-
lution.
Figure.8 shows the statistics of various param-
DEVELOPMENTAL PLASTICITY IN CARTESIAN GENETIC PROGRAMMING BASED NEURAL NETWORKS
455
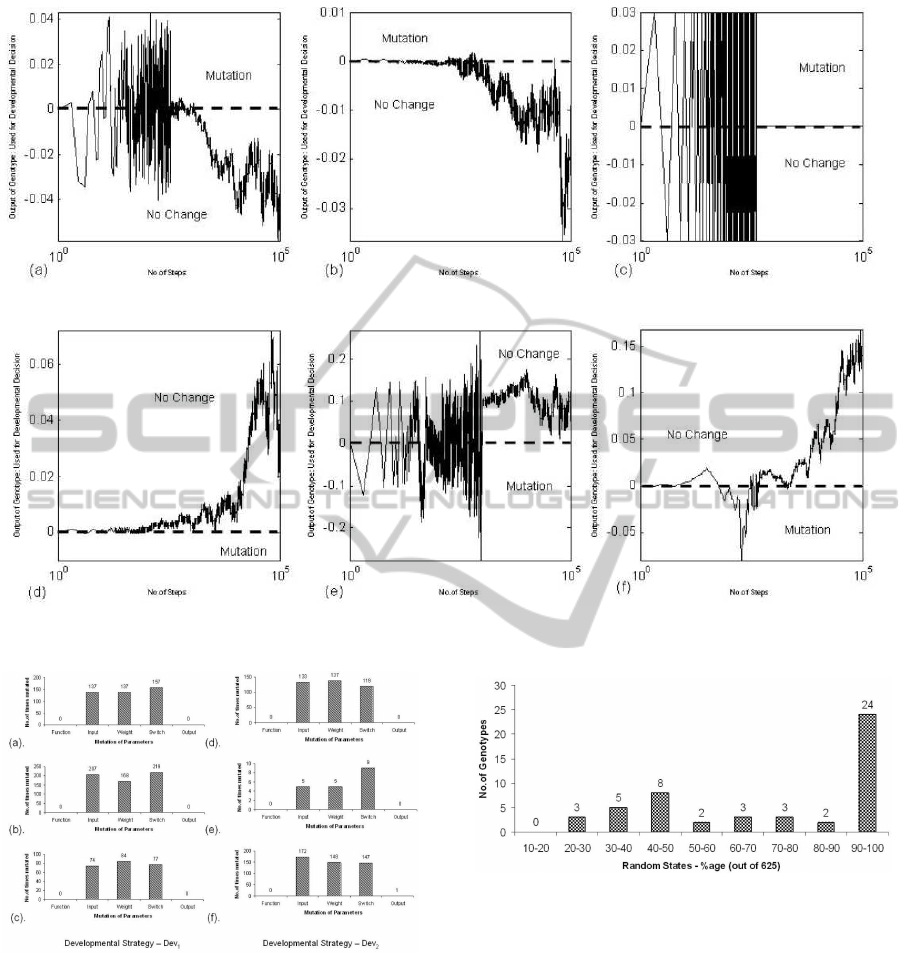
Figure 7: Developmental Behaviour of the network - (a,b,c) Dev
1
Strategy and (d,e,f) Dev
2
strategy.
Figure 8: Statistics of the changes in network parameters
for the six cases in Figure 7.
eters that are mutated for the six cases in Figure.7
during 100,000 time steps while the pole(s) are be-
ing balanced. A similar trend is observed in all the
cases with switches, inputs and weights updated with
almost same ratio. On average the network is changed
by 0.3 to 0.4% during runtime with most (95%) devel-
opment taking place at the early stage as evident from
Figure.7.
As learning to learn is a complex dynamic nonlin-
Figure 9: SinglePole Task-Number of Genotypes that are
successful in balancing the pole in 625 random initial states.
ear process even a biological brain does not process
inputs linearly. Remaining dormant for a number of
steps reduces the chance of quick learning while it is
observed that changing and adapting (to remain dor-
mant) to the change is an effective strategy of learn-
ing.
8.2 Robustness of PCGPANN
The robustness of the genotypes is inferred by sub-
jecting the genotype to 625 random initial states and
checking it for 1000 steps only. If the genotype is
able to balance then it is termed as robust. Figure.
ICINCO 2011 - 8th International Conference on Informatics in Control, Automation and Robotics
456

Figure 10: DoublePole Task-Number of Genotypes that are
successful in balancing the pole in 625 random initial states.
9 and 10 represents the performance of 50 genotypes
(solutions of the single and double pole task) on 625
random initial states. For the single pole, 48% of
the genotype were able to balance at 90-100% (which
corresponds to 563-625) of the random states which
can be inferred from the graph presented in Figure.9,
thus attaining an average of 456. While for the dou-
ble pole 44% genotypes were able to balance at 60-
100% of the random states achieving an average of
349. Thus the genotypes undergoing developmental
plasticity during the 1000 steps exhibit robustness.
Table 3: Average number of random cart-pole initializations
(out of 625) that can be solved.
Type SinglePole DoublePole
FCGPANN 590 277.38
PCGPANN 456 349
Table.3 shows the generalization of the PCG-
PANN genotypes in comparison to feedforward CG-
PANN for both the single and double pole balancing
scenarios. The PCGPANN strategy proves more ro-
bust in the double pole balancing task (which is a non-
linear problem) where on average a genotype at 349
out of 625 different initial states is developed into suc-
cessful and feasible solutions.
9 CONCLUSIONS
In this paper, plastic feedforward neural network
based on the representation of Cartesian Genetic Pro-
gramming (PCGPANN) is proposed and applied on
a standard benchmark control problems. Plastic-
ity involves mutating the neural network parameters
namely weight, switches, inputs, functions and out-
put at run-time. Plastic/Developmental decisions are
made by using a developmental output from the net-
work which induces plastic changes at runtime. The
neurocontrollers produced by PCGPANN algorithm
were found to be more robust than its FCGPANN
counterpart. The network phenotype changes during
run time, thus modifying its architecture in response
to changing input patterns. The algorithm was also
found to outperform most of the static neuroevolu-
tionary algorithms in terms of speed of learning. It
should be noted that the phenotypic mutations happen
each time the new output is obtained from the evolved
program in the genotype. However, in principle, this
could be done at a slower rate (for instance, after a
certain number of evolved program executions, rather
than at every timestep). This remains to be investi-
gated.
In future plasticity will be investigated for recur-
rent networks and tested for the non-markovian con-
trol problems. The goal is to investigate whether in-
duction of both signal based feedback (recurrent) and
structural feedback (plasticity) can enhance the per-
formance. Also we intend to examine whether the
PCGPANN is able to solve problems faster and more
accurately by obtaining repeated experienceof its task
environment (post evolution). Eventually the aim is
to investigate whether PCGPANNs can be trained to
solve a sequence of problems without forgetting how
to solve earlier problems.
REFERENCES
Ans, B., Rousset, S., French, R. M., and Musca, S.
(2002). Preventing catastrophic interference in
multiple-sequence learning using coupled reverberat-
ing elman networks. In Proc. 24th, Annual conf. of
cognitive science society, pages 71–76.
Baxter, J. (1992). The evolution of learning algorithms for
artificial neural networks. D.Green & T.Bossomaier,
Complex Systems, pages 313 – 326.
Cangelosi, A., Nolfi, S., and Parisi, D. (1994). Cell division
and migration in a ’genotype’ for neural networks.
Network-Computation in Neural Systems, 5:497–515.
Clune, J., Beckmann, B. E., Ofria, C., and Pennock,
R. T. (2008). Evolving coordinated quadruped gaits
with the hyperneat generative encoding. Proc. IEEE
CEC’2008, pages 2764–2771.
Dalaert, F. and Beer, R. (1994). Towards an evolvable
model of development for autonomous agent synthe-
sis. In Brooks, R. and Maes, P. eds. Proc. 4rth Conf.
on Artificial Life. MIT Press.
Floreano, D. and Urzelai, J. (2000). Evolutionary robots
with online self-organization and behavioral fitness.
Neural Networks, 13:431 – 443.
French, R. M. (1994). Catastrophic forgetting in connec-
tionist networks: Causes, consequences and solutions.
In Trends in Cognitive Sciences, pages 128–135.
DEVELOPMENTAL PLASTICITY IN CARTESIAN GENETIC PROGRAMMING BASED NEURAL NETWORKS
457

Gomez, F., Schmidhuber, J., and Miikkulainen, R. (2008).
Accelerated neural evolution through cooperatively
coevolved synapses. J. Mach. Learn. Res., 9:937–965.
Gomez, F. J. and Miikkulainen, R. (1999). Solving non-
markovian control tasks with neuroevolution. In Proc.
Int. joint Conf. on Artificial intelligence, pages 1356–
1361. Morgan Kaufmann Publishers Inc.
Gruau, F. (1994). Automatic definition of modular neural
networks. Adaptive Behaviour, 3:151–183.
Gruau, F., Whitley, D., and Pyeatt, L. (1996). A com-
parison between cellular encoding and direct encod-
ing for genetic neural network. In Genetic Program-
ming 1996:Proceeding of the First Annual conference,
pages 81–89 MIT Press.
Harding, S., Miller, J. F., and Benzhaf, W. (2010).
Developments in cartesian genetic program-
ming:selfmodifying cgp. GPEM, 11(2):397–439.
Hussain, T. and Browse, R. (2000). Evolving neural net-
works using attribute grammars. IEEE Symp. Combi-
nations of Evolutionary Computation and Neural Net-
works, 2000, pages 37 – 42.
Jacob, C. and Rehder, J. (1993). Evolution of neural
net architectures by a hierarchical grammar-based ge-
netic system. In Proc. ICANNGA93, pages 72–79.
Springer-Verlag.
Kandel, E. R., Schwartz, J. H., and Jessell (2002). Princi-
ples of Neural Science, 4rth Edition. McGraw-Hill.
Khan, G., Miller, J., and Halliday, D. (2007). Coevolution
of intelligent agents using cartesian genetic program-
ming. In Proc. GECCO’2007, pages 269 – 276.
Khan, M., Khan, G., and F. Miller, J. (2010a). Evolution
of optimal anns for non-linear control problems us-
ing cartesian genetic programming. In Proc. IEEE.
ICAI’2010.
Khan, M., Khan, G., and Miller, J. (2010b). Effi-
cient representation of recurrent neural networks for
markovian/non-markovian non-linear control prob-
lems. In Proc. ISDA’2010, pages 615–620.
Kitano, H. (1990). Designing neural networks using ge-
netic algorithm with graph generation system. Com-
plex Systems, 4:461–476.
McCloskey, M. and Cohen, N. (1989). Catastrophic in-
terference in connectionist networks: The sequential
learning problem. The Psychology of Learning and
Motivation, 24:109–165.
Miller, J. and Smith, S. (2006). Redundancy and com-
putation efficiency in cartesian genetic programming.
IEEE Trans. Evol. Comp., 10:167–174.
Miller, J. F. and Thomson, P. (2000). Cartesian genetic
programming. In Proc. EuroGP’2000, volume 1802,
pages 121–132.
Moriarty, D. (1997). Symbiotic Evolution of Neural Net-
works in Sequential Decision Tasks. PhD thesis, Uni-
versity of Texas at Austin.
Nicholas F.McPhee, Ellery Crane, S. E. and Poli, R. (2009).
Developmental plasticity in linear genetic program-
ming. Proc. GECCO’2009, pages 1019–1026.
Nolfi, S., Miglino, O., and Parisi, D. (1994). Phenotypic
plasticity in evolving neural networks. In Proc. Int.
Conf. from perception to action. IEEE Press.
Ratcliff, R. (1990). Connectionist models of recognition
and memory:constraints imposed by learning and for-
getting functions. Psychological Review, 97:205–308.
Risi, S., Hughes, C. E., and Stanley, K. O. (2010). Evolving
plastic neural networks with novelty search. Adaptive
Behavior.
Rivero, D., Rabual, J., Dorado, J., and Pazos, A. (2007).
Automatic design of anns by means of gp for
data mining tasks: Iris flower classification prob-
lem. Adaptive and Natural Computing Algorithms,
4431:276–285.
Roggen, D., Federici, D., and Floreano, D. (2007). Evo-
lutionary morphogenesis for multi-cellular systems.
GPEM, 8:61–96.
Rust, A., Adams, R., and H., B. (2000). Evolutionary neural
topiary: Growing and sculpting artificial neurons to
order. In Proc. ALife VII, pages 146–150. MIT Press.
Sharkey, N. and Sharkey, A. (1995). An analysis of catas-
trophic interference. Connection Science, 7(3-4):301–
330(30).
Sims, K. (1994). Evolving 3d morphology and behavior
by competition. In Artificial life 4 proceedings, pages
28–39. MIT Press.
Stanley, K. O., D’Ambrosio, D. B., and Gauci, J. (2009).
A hypercube-based encoding for evolving large-scale
neural networks. Artif. Life, 15:185–212.
Stanley, K. O. and Miikkulainen, R. (2002). Evolving neu-
ral network through augmenting topologies. Evolu-
tionary Computation, 10(2):99–127.
Wieland, A. P. (1991). Evolving neural network controllers
for unstable systems. In Proc. Int. Joint Conf. Neural
Networks, pages 667–673.
ICINCO 2011 - 8th International Conference on Informatics in Control, Automation and Robotics
458
